Вам также может понравиться
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)От EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Рейтинг: 4.5 из 5 звезд4.5/5 (121)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryОт EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryРейтинг: 3.5 из 5 звезд3.5/5 (231)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaОт EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaРейтинг: 4.5 из 5 звезд4.5/5 (266)
- Never Split the Difference: Negotiating As If Your Life Depended On ItОт EverandNever Split the Difference: Negotiating As If Your Life Depended On ItРейтинг: 4.5 из 5 звезд4.5/5 (838)
- The Emperor of All Maladies: A Biography of CancerОт EverandThe Emperor of All Maladies: A Biography of CancerРейтинг: 4.5 из 5 звезд4.5/5 (271)
- The Little Book of Hygge: Danish Secrets to Happy LivingОт EverandThe Little Book of Hygge: Danish Secrets to Happy LivingРейтинг: 3.5 из 5 звезд3.5/5 (400)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeОт EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeРейтинг: 4 из 5 звезд4/5 (5794)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyОт EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyРейтинг: 3.5 из 5 звезд3.5/5 (2259)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreОт EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreРейтинг: 4 из 5 звезд4/5 (1090)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersОт EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersРейтинг: 4.5 из 5 звезд4.5/5 (345)
- Team of Rivals: The Political Genius of Abraham LincolnОт EverandTeam of Rivals: The Political Genius of Abraham LincolnРейтинг: 4.5 из 5 звезд4.5/5 (234)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceОт EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceРейтинг: 4 из 5 звезд4/5 (895)
- The Unwinding: An Inner History of the New AmericaОт EverandThe Unwinding: An Inner History of the New AmericaРейтинг: 4 из 5 звезд4/5 (45)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureОт EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureРейтинг: 4.5 из 5 звезд4.5/5 (474)
- VC AndrewsДокумент3 страницыVC AndrewsLesa O'Leary100% (1)
- The Yellow House: A Memoir (2019 National Book Award Winner)От EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Рейтинг: 4 из 5 звезд4/5 (98)
- Current Surgical Therapy 13th EditionДокумент61 страницаCurrent Surgical Therapy 13th Editiongreg.vasquez490100% (41)
- Mindray PM 9000 User ID10240 PDFДокумент378 страницMindray PM 9000 User ID10240 PDFJuan FernandoОценок пока нет
- MOS - Steel StructureДокумент15 страницMOS - Steel StructuredennisОценок пока нет
- Barista Skills Foundation Curriculum enДокумент4 страницыBarista Skills Foundation Curriculum enCezara CarteșОценок пока нет
- CUET 2022 General Test 6th October Shift 1Документ23 страницыCUET 2022 General Test 6th October Shift 1Dhruv BhardwajОценок пока нет
- Marieb ch3dДокумент20 страницMarieb ch3dapi-229554503Оценок пока нет
- Coco Mavdi Esl5Документ6 страницCoco Mavdi Esl5gaurav222980Оценок пока нет
- Jota - EtchДокумент3 страницыJota - EtchRidwan BaharumОценок пока нет
- Atlascopco XAHS 175 DD ASL Parts ListДокумент141 страницаAtlascopco XAHS 175 DD ASL Parts ListMoataz SamiОценок пока нет
- Lotus Exige Technical InformationДокумент2 страницыLotus Exige Technical InformationDave LeyОценок пока нет
- Junos ErrorsДокумент2 страницыJunos ErrorsrashidsharafatОценок пока нет
- 2432 - Test Solutions - Tsol - 2432 - 21702Документ5 страниц2432 - Test Solutions - Tsol - 2432 - 21702Anmol PanchalОценок пока нет
- Lakh Only) Being The Amount Covered Under The Aforesaid Dishonoured Cheque, and So AlsoДокумент2 страницыLakh Only) Being The Amount Covered Under The Aforesaid Dishonoured Cheque, and So AlsoShivam MishraОценок пока нет
- Inspection Report For Apartment Building at 1080 93rd St. in Bay Harbor IslandsДокумент13 страницInspection Report For Apartment Building at 1080 93rd St. in Bay Harbor IslandsAmanda RojasОценок пока нет
- Advanced Java SlidesДокумент134 страницыAdvanced Java SlidesDeepa SubramanyamОценок пока нет
- Weg CFW500 Enc PDFДокумент32 страницыWeg CFW500 Enc PDFFabio Pedroso de Morais100% (1)
- Wins Salvacion Es 2021Документ16 страницWins Salvacion Es 2021MURILLO, FRANK JOMARI C.Оценок пока нет
- Donnan Membrane EquilibriaДокумент37 страницDonnan Membrane EquilibriamukeshОценок пока нет
- Tourbier Renewal NoticeДокумент5 страницTourbier Renewal NoticeCristina Marie DongalloОценок пока нет
- Assignment RoadДокумент14 страницAssignment RoadEsya ImanОценок пока нет
- Investigative JournalismДокумент8 страницInvestigative JournalismMitchie LlagasОценок пока нет
- Anviz T5 RFID ManualДокумент52 страницыAnviz T5 RFID ManualLuis Felipe Olaya SandovalОценок пока нет
- Durability of Prestressed Concrete StructuresДокумент12 страницDurability of Prestressed Concrete StructuresMadura JobsОценок пока нет
- R15 Understanding Business CyclesДокумент33 страницыR15 Understanding Business CyclesUmar FarooqОценок пока нет
- Module 2Документ7 страницModule 2karthik karti100% (1)
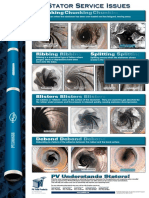
- Chunking Chunking Chunking: Stator Service IssuesДокумент1 страницаChunking Chunking Chunking: Stator Service IssuesGina Vanessa Quintero CruzОценок пока нет
- SAP HR - Legacy System Migration Workbench (LSMW)Документ5 страницSAP HR - Legacy System Migration Workbench (LSMW)Bharathk KldОценок пока нет
- SodiumBenzoate PDFДокумент3 страницыSodiumBenzoate PDFyotta024Оценок пока нет
- Ged 102 Mathematics in The Modern WorldДокумент84 страницыGed 102 Mathematics in The Modern WorldKier FormelozaОценок пока нет