Вам также может понравиться
- Guía de EstudioДокумент5 страницGuía de EstudioAnderson CuatindioyОценок пока нет
- Absorcion - Excrecion - EliminacionДокумент5 страницAbsorcion - Excrecion - EliminacionEly SuarezОценок пока нет
- Resumen Membranas BiologicasДокумент6 страницResumen Membranas BiologicaslizzyrodnuzОценок пока нет
- EnzimasДокумент16 страницEnzimasSeoba LeeОценок пока нет
- Tejido ConectivoДокумент36 страницTejido ConectivoLuZerO KaTheRinE100% (1)
- Bioquimica Metabolismo de La Prostaglandina y ColesterolДокумент9 страницBioquimica Metabolismo de La Prostaglandina y Colesterolmarce bastidasОценок пока нет
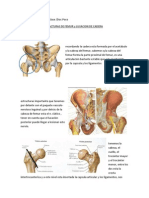
- Musculos de La Región GlúteaДокумент8 страницMusculos de La Región GlúteaMarco Diaz LeonОценок пока нет
- Interacciones Entre Celulas y Matriz ExtracelularДокумент10 страницInteracciones Entre Celulas y Matriz ExtracelularIngridОценок пока нет
- Aparato de GolgiДокумент34 страницыAparato de GolgiMoisés J. Duran CanevaroОценок пока нет
- Los MúsculosДокумент5 страницLos MúsculosrafaeltorralbapОценок пока нет
- Conceptos Basicos InmunidadДокумент9 страницConceptos Basicos InmunidadYessica FuentesОценок пока нет
- Notas de Clase Sobre La CélulaДокумент3 страницыNotas de Clase Sobre La CélulaNicoleAndreaPérezОценок пока нет
- Ciclo de Krebs - Exposición de F. de Micro y BioДокумент29 страницCiclo de Krebs - Exposición de F. de Micro y BioMaria Jose QuirozОценок пока нет
- Repaso. Fisiologia RespiratoriaДокумент36 страницRepaso. Fisiologia Respiratoriamarisel ruizОценок пока нет
- Monografia HistologiaДокумент29 страницMonografia HistologiaMario Lazo100% (1)
- Membrana CelularДокумент35 страницMembrana CelularCoral PeñaОценок пока нет
- Trabajo de Liquidos CorporalesДокумент8 страницTrabajo de Liquidos Corporalesharas_18_83Оценок пока нет
- Test de Histología Primero MCMДокумент1 страницаTest de Histología Primero MCMProf. Víctor M. VitoriaОценок пока нет
- Contenido Unidades de Catedra Anatomia UcvДокумент37 страницContenido Unidades de Catedra Anatomia UcvLuis Manuel Acosta MarcanoОценок пока нет
- Qué Es El HidrosalpinxДокумент3 страницыQué Es El HidrosalpinxEvelin UribeОценок пока нет
- Microvasculatura Del Tubo DigestivoДокумент3 страницыMicrovasculatura Del Tubo DigestivoClaudioVillalobosОценок пока нет
- Histologia Aparato DigestivoДокумент9 страницHistologia Aparato DigestivoDayana CoyagoОценок пока нет
- Transporte Activo y PasivoДокумент9 страницTransporte Activo y PasivoDavid Marin Antonio100% (2)
- Enzimas AmilolíticasДокумент2 страницыEnzimas AmilolíticasJonathan100% (1)
- Mitocondria y VacuolaДокумент22 страницыMitocondria y VacuolaDagoberto Durán100% (1)
- Metaplasia Escamosa Clase CitoДокумент23 страницыMetaplasia Escamosa Clase Citopedro_gomarОценок пока нет
- Sobre Sistema LinfaticoДокумент6 страницSobre Sistema LinfaticoSaul Guevara0% (1)
- Semana 05 EyF - Guia de Práctica 05Документ9 страницSemana 05 EyF - Guia de Práctica 05carlosbullonperuОценок пока нет
- Reticulo Endoplasmatico Liso (Rel)Документ14 страницReticulo Endoplasmatico Liso (Rel)Yeison PelufoОценок пока нет
- Datos Claves de AnatomíaДокумент48 страницDatos Claves de AnatomíaGuerrero Omar0% (1)
- 2f 3c-Fracturas de Femur Luxacion de CaderaДокумент26 страниц2f 3c-Fracturas de Femur Luxacion de CaderaasdgiaybhiОценок пока нет
- Tejido Epitelial Glandular. Mapa ConceptualДокумент3 страницыTejido Epitelial Glandular. Mapa ConceptualGretel TroetschОценок пока нет
- Capitulo 27 StryerДокумент21 страницаCapitulo 27 StryerCristobal Ibarra SalazarОценок пока нет
- Informe Histologia Baez 2 Semestre Nº4Документ6 страницInforme Histologia Baez 2 Semestre Nº4Simoné Pazmiño GranadaОценок пока нет
- BIOFÍSICA DEL SISTEMA RENAL ChidoooДокумент52 страницыBIOFÍSICA DEL SISTEMA RENAL ChidoooIsidro Medina100% (1)
- Resumen Tejido Conjuntivo o ConectivoДокумент2 страницыResumen Tejido Conjuntivo o ConectivoOrlandoArtemioPoloVeraОценок пока нет
- El BazoДокумент2 страницыEl BazoAngela Maria SandovalОценок пока нет
- Guía de Estudio 6Документ8 страницGuía de Estudio 6Cce ReginaОценок пока нет
- Tejido AdiposoДокумент24 страницыTejido AdiposoEdu Sama Murillo MirandaОценок пока нет
- Apunte Sistema Circulatorio PDFДокумент16 страницApunte Sistema Circulatorio PDFMabel Elisa Rodriguez MendezОценок пока нет
- Sistema Respiratorio Morfo IVДокумент2 страницыSistema Respiratorio Morfo IVKarla M. FrancoОценок пока нет
- Procesos de GestaciónДокумент67 страницProcesos de Gestaciónleondario223hotmailcОценок пока нет
- Tejido MuscularДокумент20 страницTejido MuscularIdalia GermánОценок пока нет
- Tema 1 Células y Sus FuncionesДокумент16 страницTema 1 Células y Sus Funcioneselias ayala100% (1)
- Bioq 6Документ3 страницыBioq 6Luis Arevalo100% (1)
- Transporte CelularДокумент12 страницTransporte CelularFelipeMendozaОценок пока нет
- Aminopeptidasa PDFДокумент14 страницAminopeptidasa PDFNathalia Montaño100% (1)
- Generalidades de Los TejidosДокумент6 страницGeneralidades de Los TejidosJuan Antonio100% (2)
- Diapositivas ExposiciónДокумент33 страницыDiapositivas ExposiciónNataly PillajoОценок пока нет
- Cap 17Документ27 страницCap 17Dulce MedinaОценок пока нет
- La CelulaДокумент4 страницыLa CelulamiguelОценок пока нет
- Balsas LipidicasДокумент5 страницBalsas LipidicasJuan Jesus Barrios CapuchinoОценок пока нет
- Discusión N - 1 de Bioquímica IДокумент69 страницDiscusión N - 1 de Bioquímica IVictor Rivera75% (4)
- Guia Practica - Sesión 6 - Sistema Esquelético IДокумент6 страницGuia Practica - Sesión 6 - Sistema Esquelético ILizbeth Rodriguez PorrasОценок пока нет
- MITOCONDRIAДокумент7 страницMITOCONDRIAJorge Luis100% (1)
- Funciones Del AmpcДокумент3 страницыFunciones Del AmpcRaulОценок пока нет
- Biología Sección 4Документ13 страницBiología Sección 4Choque Condori NoeliaОценок пока нет
- Las Membranas de La Célula Ejercen Diversas ActividadesДокумент13 страницLas Membranas de La Célula Ejercen Diversas ActividadesKauely SouzaОценок пока нет
- Apuntes Clases Ene Jun 2019Документ121 страницаApuntes Clases Ene Jun 2019Kimberly MartinezОценок пока нет
- Estructura de La MembranaДокумент15 страницEstructura de La MembranaSu SaucedoОценок пока нет
- Qué Es Educación para La PazДокумент9 страницQué Es Educación para La PazhenzejulyОценок пока нет
- Enfermedades Psicosomáticas e Histeria de ConversiónДокумент5 страницEnfermedades Psicosomáticas e Histeria de ConversiónhenzejulyОценок пока нет
- ArbolДокумент5 страницArbolhenzejulyОценок пока нет
- El Cerebro en Los Zurdos - Karen Maribel Soto MercadoДокумент6 страницEl Cerebro en Los Zurdos - Karen Maribel Soto MercadohenzejulyОценок пока нет
- Tortugas Hasta El Fondo - Chapter 1Документ84 страницыTortugas Hasta El Fondo - Chapter 1henzejulyОценок пока нет
- Monet ClaudeДокумент150 страницMonet ClaudehenzejulyОценок пока нет
- La HojaДокумент5 страницLa HojahenzejulyОценок пока нет
- Materiales A Revisar Comunidad InfantilДокумент1 страницаMateriales A Revisar Comunidad InfantilhenzejulyОценок пока нет
- Desviaciones PsiquicasДокумент18 страницDesviaciones PsiquicashenzejulyОценок пока нет
- ADAPTACIÓNДокумент3 страницыADAPTACIÓNhenzejulyОценок пока нет
- Cuadro ReinosДокумент2 страницыCuadro ReinoshenzejulyОценок пока нет
- Guia de ObservacionДокумент14 страницGuia de ObservacionhenzejulyОценок пока нет
- Túnica Nerviosa o Retina 11Документ34 страницыTúnica Nerviosa o Retina 11henzejuly50% (2)
- Usos de La EpidemiologiaДокумент23 страницыUsos de La EpidemiologiahenzejulyОценок пока нет
- Características Ultraestructurales y Morfología de Los VirusДокумент26 страницCaracterísticas Ultraestructurales y Morfología de Los VirusNayely Miguel100% (1)
- Reticulo Endoplasmatico LisoДокумент7 страницReticulo Endoplasmatico LisoWilber de la CruzОценок пока нет
- Cuadro Comparativo Entre Las Similitudes y Diferencias Entre Las Células Procariotas y EucariotasДокумент6 страницCuadro Comparativo Entre Las Similitudes y Diferencias Entre Las Células Procariotas y Eucariotasjenny patricia carabaliОценок пока нет
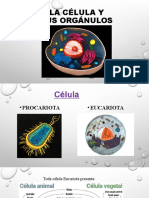
- La Célula y Sus OrgánulosДокумент35 страницLa Célula y Sus OrgánulosQuimica GonzalezОценок пока нет
- Clase 7 de BiologiaДокумент15 страницClase 7 de BiologiaJose RamirezОценок пока нет
- Ponencia-Ascenso - Ciencia y AmbienteДокумент80 страницPonencia-Ascenso - Ciencia y AmbienteJesus Alcocer100% (1)
- Celulas Vegetal y Animal (Cuadro Comparativo)Документ17 страницCelulas Vegetal y Animal (Cuadro Comparativo)bercelisОценок пока нет
- Concepto de Celula, Linea de Tiempo Celula Eucariota y ProcariotaДокумент15 страницConcepto de Celula, Linea de Tiempo Celula Eucariota y ProcariotaYajaira ReyesОценок пока нет
- La Célula 10Документ24 страницыLa Célula 10Quimica SebastianaОценок пока нет
- La Celula ProcariotaДокумент3 страницыLa Celula ProcariotaMiguel Ruiz MartínezОценок пока нет
- Evaluación Sumativa CelulaДокумент3 страницыEvaluación Sumativa Celulaescuela chilecito100% (1)
- Dsllo Módulo 2 - Ciencias NaturalesДокумент21 страницаDsllo Módulo 2 - Ciencias NaturalesIsabella Sierra67% (3)
- Estructura de Los Seres VivosДокумент3 страницыEstructura de Los Seres Vivospcastillo_681263Оценок пока нет
- Resumen Citología, Estructura y Función CelularДокумент2 страницыResumen Citología, Estructura y Función CelularKatherine Beraún60% (5)
- Prueba Octavo CèlulaДокумент4 страницыPrueba Octavo CèlulaAlvarez MelissaОценок пока нет
- La Diferenciación CelularДокумент2 страницыLa Diferenciación CelularLourdes RojoОценок пока нет
- Biologia CelularДокумент1 страницаBiologia CelularAnnevelis Agreda SalazarОценок пока нет
- Team With NutrientsДокумент125 страницTeam With Nutrientsleandro19922100% (1)
- Morfología Humana. Tomo 1Документ199 страницMorfología Humana. Tomo 1Jose Alberto Andrade López90% (41)
- MA526 - S03 - S06 - Lectura 02 - La CélulaДокумент24 страницыMA526 - S03 - S06 - Lectura 02 - La Célulamendoza sanchizОценок пока нет
- Naturales 9 Modulo 1Документ16 страницNaturales 9 Modulo 1Angel Ortiz CОценок пока нет
- La CélulaДокумент21 страницаLa CélulaAndrés E. Espinoza NolascoОценок пока нет
- Normalidad de Celulas EpitelialesДокумент5 страницNormalidad de Celulas EpitelialesPulido3108Оценок пока нет
- Ciencias Clei 3 PDFДокумент61 страницаCiencias Clei 3 PDFSILVIA YANETH MORA RAMIREZОценок пока нет
- Trabajo Ex Aula Situación CovidДокумент48 страницTrabajo Ex Aula Situación CovidJared HernandezОценок пока нет
- Triptico Celula AnimalДокумент2 страницыTriptico Celula AnimalOscar Ccoycca Fernandez82% (11)
- Guia Laboratorio PRÁCTICA No 3Документ10 страницGuia Laboratorio PRÁCTICA No 3Alejandra MolinaОценок пока нет
- Observación de Células de Epidermis de Cebolla InformeДокумент8 страницObservación de Células de Epidermis de Cebolla InformeMARTA JULIÁN PASTORОценок пока нет
- Organelos ClasificacionДокумент3 страницыOrganelos ClasificacionMitzy Saucedo33% (3)