Вам также может понравиться
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceОт EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceРейтинг: 4 из 5 звезд4/5 (895)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeОт EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeРейтинг: 4 из 5 звезд4/5 (5794)
- Are SH-SY5Y and MN9D Cell Lines Truly Dopaminergic?: Inoka S. Balasooriya and Kandatage WimalasenaДокумент2 страницыAre SH-SY5Y and MN9D Cell Lines Truly Dopaminergic?: Inoka S. Balasooriya and Kandatage WimalasenaHosam AlbОценок пока нет
- A Comparison of Protein Extraction Methods Using Organic Solvents For Secretome of Aspergillus Fumigatus Strain (MTCC 1811)Документ2 страницыA Comparison of Protein Extraction Methods Using Organic Solvents For Secretome of Aspergillus Fumigatus Strain (MTCC 1811)Hosam AlbОценок пока нет
- Comparison of Three Routine Protein Precipitation Methods: Acetone, TCA/acetone Wash and TCA/acetoneДокумент3 страницыComparison of Three Routine Protein Precipitation Methods: Acetone, TCA/acetone Wash and TCA/acetoneHosam AlbОценок пока нет
- Drug Development ProcessДокумент1 страницаDrug Development ProcessHosam AlbОценок пока нет
- The Yellow House: A Memoir (2019 National Book Award Winner)От EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Рейтинг: 4 из 5 звезд4/5 (98)
- The Little Book of Hygge: Danish Secrets to Happy LivingОт EverandThe Little Book of Hygge: Danish Secrets to Happy LivingРейтинг: 3.5 из 5 звезд3.5/5 (400)
- Never Split the Difference: Negotiating As If Your Life Depended On ItОт EverandNever Split the Difference: Negotiating As If Your Life Depended On ItРейтинг: 4.5 из 5 звезд4.5/5 (838)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureОт EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureРейтинг: 4.5 из 5 звезд4.5/5 (474)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryОт EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryРейтинг: 3.5 из 5 звезд3.5/5 (231)
- The Emperor of All Maladies: A Biography of CancerОт EverandThe Emperor of All Maladies: A Biography of CancerРейтинг: 4.5 из 5 звезд4.5/5 (271)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaОт EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaРейтинг: 4.5 из 5 звезд4.5/5 (266)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersОт EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersРейтинг: 4.5 из 5 звезд4.5/5 (345)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyОт EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyРейтинг: 3.5 из 5 звезд3.5/5 (2259)
- Team of Rivals: The Political Genius of Abraham LincolnОт EverandTeam of Rivals: The Political Genius of Abraham LincolnРейтинг: 4.5 из 5 звезд4.5/5 (234)
- The Unwinding: An Inner History of the New AmericaОт EverandThe Unwinding: An Inner History of the New AmericaРейтинг: 4 из 5 звезд4/5 (45)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreОт EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreРейтинг: 4 из 5 звезд4/5 (1090)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)От EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Рейтинг: 4.5 из 5 звезд4.5/5 (121)
- Chapt 27 Ap2Документ5 страницChapt 27 Ap2candybio83Оценок пока нет
- Cranial Nerve: Dr. Dian Prasetyo Wibisono, M.SCДокумент49 страницCranial Nerve: Dr. Dian Prasetyo Wibisono, M.SCgeneОценок пока нет
- 5096 w07 Ms 2Документ6 страниц5096 w07 Ms 2mstudy123456Оценок пока нет
- Syndrome Differentiation of The Zang-Fu Organs - 1Документ99 страницSyndrome Differentiation of The Zang-Fu Organs - 1clerise3Оценок пока нет
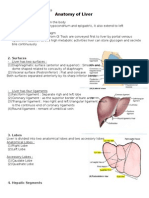
- Anatomy of LiverДокумент2 страницыAnatomy of LiverelsafirdaaaОценок пока нет
- Heart Diagram: Name: - DateДокумент3 страницыHeart Diagram: Name: - DateTheresa GarrettОценок пока нет
- Classes of Vertebrates (Comparative Analysis)Документ173 страницыClasses of Vertebrates (Comparative Analysis)Vinci EbordeОценок пока нет
- 2Документ2 страницы2test1245 testОценок пока нет
- Haemostasis: Case StudyДокумент19 страницHaemostasis: Case StudyTusabe FredОценок пока нет
- Test Bank For Radiographic Pathology For Technologists 7th Edition by KowalczykДокумент8 страницTest Bank For Radiographic Pathology For Technologists 7th Edition by KowalczykAudrey Snook100% (31)
- Body Lesson 1Документ10 страницBody Lesson 1Vadim BulbakОценок пока нет
- BPT First Year Syllabus Rabindranath Tagore UniversityДокумент33 страницыBPT First Year Syllabus Rabindranath Tagore Universitysubhu7678Оценок пока нет
- 02 IVD Hernia Case Report MedBac&IEДокумент18 страниц02 IVD Hernia Case Report MedBac&IEClaude ReyesОценок пока нет
- BBДокумент141 страницаBBbhimaОценок пока нет
- Endocrine System Worksheet AnswersДокумент3 страницыEndocrine System Worksheet AnswersmariaОценок пока нет
- Exercise 3 Fixation StudentДокумент7 страницExercise 3 Fixation StudentErika Nicole SiasonОценок пока нет
- BB - RH BLOOD GROUP SYSTEMДокумент5 страницBB - RH BLOOD GROUP SYSTEMWayne VillalunaОценок пока нет
- How To Dissect RatДокумент30 страницHow To Dissect RatHend MaarofОценок пока нет
- Normal ChestДокумент82 страницыNormal ChestMonica CherladyОценок пока нет
- Pridevi PREPravkeДокумент8 страницPridevi PREPravkeLukaОценок пока нет
- A Case Study of Splenic Hemangiosarcoma in A Bitch and Its Surgical ManagementДокумент3 страницыA Case Study of Splenic Hemangiosarcoma in A Bitch and Its Surgical ManagementWira KusumaОценок пока нет
- The Allergic Response: Kathleen Nicole B. Bidua First Year ResidentДокумент44 страницыThe Allergic Response: Kathleen Nicole B. Bidua First Year ResidentNicole BiduaОценок пока нет
- 2021 Temporal Bone Imaging-Made-EasyДокумент152 страницы2021 Temporal Bone Imaging-Made-EasyJaime RimoldiОценок пока нет
- 4 ANA212 WORKSHEET FoundTopicДокумент8 страниц4 ANA212 WORKSHEET FoundTopickirsten merielleОценок пока нет
- Group 1-BSN 1-N MC1 AnaphyДокумент4 страницыGroup 1-BSN 1-N MC1 AnaphyCarla Diana BuenacosaОценок пока нет
- Maslow and Collinge - Course of The Femoral Artery in The Mid - and Distal Thigh and Implications For Medial Approaches To The Distal Femur (2019)Документ5 страницMaslow and Collinge - Course of The Femoral Artery in The Mid - and Distal Thigh and Implications For Medial Approaches To The Distal Femur (2019)Kristijonas PetkevičiusОценок пока нет
- Coordinated Functions of The Nervous, Endocrine, and Reproductive SystemsДокумент30 страницCoordinated Functions of The Nervous, Endocrine, and Reproductive SystemsSarah CruzОценок пока нет
- Nursing Care Management of Patients With Anemia - 01Документ38 страницNursing Care Management of Patients With Anemia - 01Sionur 00Оценок пока нет
- 24 Fun Facts About The HeartДокумент1 страница24 Fun Facts About The HeartAljean BachinichaОценок пока нет
- Nursing Care of A Child With FractureДокумент40 страницNursing Care of A Child With FractureSomyee Pachuau100% (2)