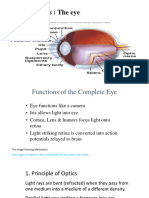
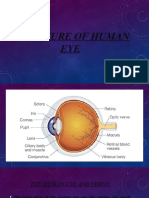
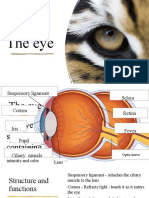
Вам также может понравиться
- 3.2 Sense OrganДокумент8 страниц3.2 Sense OrganMod HollОценок пока нет
- Biology Eye NotesДокумент12 страницBiology Eye NotesBalakrishnan MarappanОценок пока нет
- The EyeДокумент7 страницThe EyeTavvia HenryОценок пока нет
- HME 2209-wk - 2Документ7 страницHME 2209-wk - 2frank kipkoech bettОценок пока нет
- Eye Anatomy Cornea Lens: How Does Eye Accommodation Work?Документ5 страницEye Anatomy Cornea Lens: How Does Eye Accommodation Work?Trisha PersaudОценок пока нет
- Human Eye PDF 2Документ27 страницHuman Eye PDF 2Satish singhОценок пока нет
- Physics U2Документ6 страницPhysics U2Alima IqbalОценок пока нет
- The Eye and VisionДокумент70 страницThe Eye and VisionLedkos SLОценок пока нет
- The Main Parts of The Eye and Their FunctionДокумент5 страницThe Main Parts of The Eye and Their FunctionJoshuaОценок пока нет
- Human Eye and Colourful WorldДокумент11 страницHuman Eye and Colourful WorldSahishna SurapaneniОценок пока нет
- 11.human Eye and Colourful WorldДокумент22 страницы11.human Eye and Colourful WorldMalancha high school HSОценок пока нет
- Defects in Human EyeДокумент10 страницDefects in Human Eyechayan sarbadhikariОценок пока нет
- Anatomy of EyeДокумент11 страницAnatomy of EyeAbi Nan ThanОценок пока нет
- Unit 2 Windows of KnowledgeДокумент9 страницUnit 2 Windows of Knowledgesajith kumar VariathОценок пока нет
- The Human Eye and The Colourful World: Anchli Singh, Class XAДокумент17 страницThe Human Eye and The Colourful World: Anchli Singh, Class XAshashank100% (1)
- The EYE NotesДокумент6 страницThe EYE NotesSchool ApplicationОценок пока нет
- Structure and Function of The Human EyeДокумент6 страницStructure and Function of The Human EyeAljhon DelfinОценок пока нет
- 2-Special SensesДокумент63 страницы2-Special Sensestmqt2fbnzgОценок пока нет
- Physiology of VisionДокумент49 страницPhysiology of VisionPrakash PanthiОценок пока нет
- Human Eye and The Colorful World PDFДокумент12 страницHuman Eye and The Colorful World PDFJyothi DeepakОценок пока нет
- Sec 8 BДокумент3 страницыSec 8 BShiela Marie RodrigoОценок пока нет
- Anatomy Eye ReportДокумент1 страницаAnatomy Eye ReportbrylleОценок пока нет
- Physics Class X For 2017 18Документ22 страницыPhysics Class X For 2017 18KalaiОценок пока нет
- HSB Form 5 The EyeДокумент9 страницHSB Form 5 The Eyekevin maheshОценок пока нет
- 2.07 Tutorial Week 3Документ4 страницы2.07 Tutorial Week 3GraciaОценок пока нет
- 11 Human Eye and Colorful World NotesДокумент9 страниц11 Human Eye and Colorful World NotesvenkatОценок пока нет
- OLD Human Eye and The Colourful WorldДокумент11 страницOLD Human Eye and The Colourful WorldRadhika GargОценок пока нет
- CH 11Документ30 страницCH 11piyushОценок пока нет
- Eye Disorders Bio ProjectДокумент9 страницEye Disorders Bio Projectmadhav laningiriОценок пока нет
- Light-The Human and The Colourful WorldДокумент11 страницLight-The Human and The Colourful WorldTapas BanerjeeОценок пока нет
- CL X Physics Ch-11 The Human EyeДокумент12 страницCL X Physics Ch-11 The Human EyeDeepanshu BindalОценок пока нет
- 0 OphthalmologyДокумент12 страниц0 OphthalmologyParsaant SinghОценок пока нет
- Jesc111 MinДокумент12 страницJesc111 Min2021abhikchowdhuryОценок пока нет
- NCERT Books For Class 10 Science Chapter 11 Human Eye and Colorful WorldДокумент18 страницNCERT Books For Class 10 Science Chapter 11 Human Eye and Colorful WorldVanitha raoОценок пока нет
- Structure of Human Eye DipiyДокумент10 страницStructure of Human Eye Dipiyvineet_knwrОценок пока нет
- 02A Lesson Proper For Week 17: 4.1 The EyeДокумент4 страницы02A Lesson Proper For Week 17: 4.1 The EyeNanzkie Andrei SamanОценок пока нет
- WPR 2Документ5 страницWPR 2ishika mohanОценок пока нет
- 187 RefractionДокумент5 страниц187 RefractionShabir KhanОценок пока нет
- 11.human Eye and Colourful WorldДокумент19 страниц11.human Eye and Colourful WorldJagpreet SinghОценок пока нет
- 13.2 Sense Organ - EyeДокумент28 страниц13.2 Sense Organ - EyeMuhammad Amin SuhaimiОценок пока нет
- Light & Eyes: - Vision Begins When Light Comes Into The Eye - LightДокумент31 страницаLight & Eyes: - Vision Begins When Light Comes Into The Eye - LightfouadОценок пока нет
- The Human EyeДокумент3 страницыThe Human EyeKajetano GraucosОценок пока нет
- Eye Optics and AccommodationДокумент14 страницEye Optics and AccommodationJúnior AlvesОценок пока нет
- Notes Human EyeДокумент9 страницNotes Human EyeGeetanshi OberoiОценок пока нет
- HUMAN EYE Class XДокумент25 страницHUMAN EYE Class Xsai projectОценок пока нет
- Chapter 11 - Human Eye and Colourful WorldДокумент3 страницыChapter 11 - Human Eye and Colourful WorldSahil AnsariОценок пока нет
- Eye Anatomy - Review 6Документ7 страницEye Anatomy - Review 6Vidini Kusuma AjiОценок пока нет
- Chapter 11 - Human Eye and Colourful World Revision NotesДокумент3 страницыChapter 11 - Human Eye and Colourful World Revision NotesNancy GiriОценок пока нет
- CSS Everyday Science Notes PDFДокумент50 страницCSS Everyday Science Notes PDFWaqas Gul100% (1)
- Lesson 2 The Structure and Functions of The EyeДокумент12 страницLesson 2 The Structure and Functions of The EyeNguyễn Gia LộcОценок пока нет
- Notes of 10th Cbse Human EyeДокумент7 страницNotes of 10th Cbse Human EyeAvichal TatuОценок пока нет
- The Human EyeДокумент9 страницThe Human Eyegiana 4e100% (1)
- Eye IgcseДокумент4 страницыEye IgcseLavanya TheviОценок пока нет
- Human Eye PDFДокумент12 страницHuman Eye PDFMothi KarunaОценок пока нет
- Physics ProjectДокумент9 страницPhysics ProjectMedha N RaoОценок пока нет
- Lens and The EyeДокумент58 страницLens and The EyeEisle Keith Rivera TapiaОценок пока нет
- Eye and VisionДокумент20 страницEye and VisionshyanОценок пока нет
- Low Vision: Assessment and Educational Needs: A Guide to Teachers and ParentsОт EverandLow Vision: Assessment and Educational Needs: A Guide to Teachers and ParentsОценок пока нет
- SASK 105.6-Prospekt - EN - 04-2017Документ1 страницаSASK 105.6-Prospekt - EN - 04-2017Filipe MonteiroОценок пока нет
- Main Specifications of PENTAX KДокумент5 страницMain Specifications of PENTAX KVBUPDF100% (2)
- SadhnaДокумент48 страницSadhnaKumar Sumit NikhilОценок пока нет
- The Technique of Film AnimationДокумент353 страницыThe Technique of Film Animationsergey100% (1)
- Multimedia ElementsДокумент13 страницMultimedia ElementsRoderickParboTiriОценок пока нет
- Communications World 1976 Fall WinterДокумент100 страницCommunications World 1976 Fall WinterBenjamin DoverОценок пока нет
- DL004U-P Cirrus User ManualДокумент38 страницDL004U-P Cirrus User ManualJuan Pablo HernándezОценок пока нет
- DIP AssignmentДокумент2 страницыDIP AssignmentKartikeya ChauhanОценок пока нет
- FilmmakingДокумент70 страницFilmmakingKunal Chakraborty100% (4)
- Gap-Fill MCQS: The Museum of ChildhoodДокумент4 страницыGap-Fill MCQS: The Museum of ChildhoodenglishclubuОценок пока нет
- PhotojournalismДокумент18 страницPhotojournalismKook Tae HyungОценок пока нет
- Rahsul Shaw ResumeДокумент1 страницаRahsul Shaw ResumeDevin Jai-ElleОценок пока нет
- Multimedia VideoДокумент69 страницMultimedia Videoshilpashree100% (2)
- Truman ShowДокумент8 страницTruman ShowRahul GabdaОценок пока нет
- 35 Years of Murat BasdoganДокумент112 страниц35 Years of Murat BasdoganRuya Elise Peltek100% (1)
- Marketing Management: Wedding PlannersДокумент20 страницMarketing Management: Wedding Plannersscribd1314Оценок пока нет
- Visa ProceduresДокумент242 страницыVisa Proceduresjayachandran1983100% (1)
- V Ray - ManualДокумент361 страницаV Ray - Manualthanhson1985Оценок пока нет
- Bigelow v. RKO Radio Pictures, Inc., 327 U.S. 251 (1946)Документ12 страницBigelow v. RKO Radio Pictures, Inc., 327 U.S. 251 (1946)Scribd Government DocsОценок пока нет
- BRAZYLIA - Przewodnik ENG PDFДокумент21 страницаBRAZYLIA - Przewodnik ENG PDFGustavo Torrezan0% (1)
- MT6227 SchematicsДокумент7 страницMT6227 SchematicsRenan Birck PinheiroОценок пока нет
- A Basic Introduction To CCTVДокумент18 страницA Basic Introduction To CCTVsam1960100% (1)
- Lesson Plan Doc Middle Upper Secondary SextingДокумент20 страницLesson Plan Doc Middle Upper Secondary Sextingapi-475088791Оценок пока нет
- Laser Head g6f 25-28-200 Qbh100 CVWДокумент1 страницаLaser Head g6f 25-28-200 Qbh100 CVWXuan Phuong HuynhОценок пока нет
- Colour Grading - Art or ScienceДокумент44 страницыColour Grading - Art or ScienceEmile M. SmithОценок пока нет
- Medium Fonnat CamerasДокумент20 страницMedium Fonnat CamerasavhajОценок пока нет
- Edward Scissorhand'sДокумент23 страницыEdward Scissorhand'sKristie NgОценок пока нет
- Desert Magazine 1963 AprilДокумент44 страницыDesert Magazine 1963 Aprildm1937Оценок пока нет
- Module 3-Answer KeyДокумент15 страницModule 3-Answer Keyarmykrs_100% (2)
- GLOBAL Toolkit US PhaseEleven June1.2022Документ217 страницGLOBAL Toolkit US PhaseEleven June1.2022shannynnskОценок пока нет