Вам также может понравиться
- Install AquaCrop Plug-In program v6Документ2 страницыInstall AquaCrop Plug-In program v6Hector PabonОценок пока нет
- Guidelines For Installing Aquacrop: Aquacrop64Bitv61Nr02052018.ZipДокумент1 страницаGuidelines For Installing Aquacrop: Aquacrop64Bitv61Nr02052018.ZipHector PabonОценок пока нет
- Intermediate A 15cm (6 Inch) Square Makes A 7.5cm (3Документ3 страницыIntermediate A 15cm (6 Inch) Square Makes A 7.5cm (3Firoz KhanОценок пока нет
- Origami - TotoroДокумент2 страницыOrigami - Totoroapi-3709851Оценок пока нет
- Digital Origami Modeling Planar Folding PDFДокумент5 страницDigital Origami Modeling Planar Folding PDFHector PabonОценок пока нет
- Panagopoulos 2009 Visual Landscape - MNGMT & Impact AsseДокумент8 страницPanagopoulos 2009 Visual Landscape - MNGMT & Impact AsseHector PabonОценок пока нет
- Origami WolfДокумент6 страницOrigami WolfHector PabonОценок пока нет
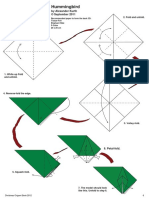
- Alexander Kurth - Hummingbird PDFДокумент9 страницAlexander Kurth - Hummingbird PDFCoolCheap ThingsОценок пока нет
- Polarbear PDFДокумент3 страницыPolarbear PDFProfe LobosОценок пока нет
- Perry Bailey - Squirrel PDFДокумент6 страницPerry Bailey - Squirrel PDFHector PabonОценок пока нет
- Intermediate A 15cm (6 Inch) Square Makes A 7.5cm (3Документ3 страницыIntermediate A 15cm (6 Inch) Square Makes A 7.5cm (3Firoz KhanОценок пока нет
- Intermediate A 15cm (6 Inch) Square Makes A 7.5cm (3Документ3 страницыIntermediate A 15cm (6 Inch) Square Makes A 7.5cm (3Firoz KhanОценок пока нет
- Ecologica Indicators 2018 Editorial-BoardДокумент1 страницаEcologica Indicators 2018 Editorial-BoardHector PabonОценок пока нет
- Ward Landcsape Scenic Quality Texas GISДокумент5 страницWard Landcsape Scenic Quality Texas GISHector PabonОценок пока нет
- Species FORMAT LIST PDFДокумент1 страницаSpecies FORMAT LIST PDFHector PabonОценок пока нет
- The Earth Care Manual GuidebookДокумент3 страницыThe Earth Care Manual GuidebookHector PabonОценок пока нет
- HUA Permaculture Lecture LinksДокумент1 страницаHUA Permaculture Lecture LinksHector PabonОценок пока нет
- EdibleДокумент11 страницEdiblecrazycarabaoОценок пока нет
- Priem 1989Документ10 страницPriem 1989Hector PabonОценок пока нет
- Rethinking Theatre of the Absurd for the EnvironmentДокумент10 страницRethinking Theatre of the Absurd for the EnvironmentHector PabonОценок пока нет
- (BBOP) - 2012. Biodiversity Offset Design Handbook-Updated PDFДокумент112 страниц(BBOP) - 2012. Biodiversity Offset Design Handbook-Updated PDFHector PabonОценок пока нет
- Frontiers in Ecology and The Environment - IssueДокумент6 страницFrontiers in Ecology and The Environment - IssueHector PabonОценок пока нет
- A Brief Tutorial On Maxent: Getting StartedДокумент39 страницA Brief Tutorial On Maxent: Getting StartedGuilherme DemetrioОценок пока нет
- Uso de La MethadonaДокумент40 страницUso de La MethadonaHector PabonОценок пока нет
- Acceptance Studies in The Field of Land Use A Critical and Syste 2018 Land UДокумент11 страницAcceptance Studies in The Field of Land Use A Critical and Syste 2018 Land UHector PabonОценок пока нет
- 3D Origami PDFДокумент94 страницы3D Origami PDFHector PabonОценок пока нет
- Biodiversity HotspotsДокумент13 страницBiodiversity HotspotsHector PabonОценок пока нет
- ColoredPencil FreemiumДокумент8 страницColoredPencil FreemiumgroventoОценок пока нет
- Carnero (00726)Документ1 страницаCarnero (00726)Hector PabonОценок пока нет
- Buitre (00140)Документ3 страницыBuitre (00140)Hector PabonОценок пока нет
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeОт EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeРейтинг: 4 из 5 звезд4/5 (5794)
- The Little Book of Hygge: Danish Secrets to Happy LivingОт EverandThe Little Book of Hygge: Danish Secrets to Happy LivingРейтинг: 3.5 из 5 звезд3.5/5 (399)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryОт EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryРейтинг: 3.5 из 5 звезд3.5/5 (231)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceОт EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceРейтинг: 4 из 5 звезд4/5 (894)
- The Yellow House: A Memoir (2019 National Book Award Winner)От EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Рейтинг: 4 из 5 звезд4/5 (98)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureОт EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureРейтинг: 4.5 из 5 звезд4.5/5 (474)
- Never Split the Difference: Negotiating As If Your Life Depended On ItОт EverandNever Split the Difference: Negotiating As If Your Life Depended On ItРейтинг: 4.5 из 5 звезд4.5/5 (838)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaОт EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaРейтинг: 4.5 из 5 звезд4.5/5 (265)
- The Emperor of All Maladies: A Biography of CancerОт EverandThe Emperor of All Maladies: A Biography of CancerРейтинг: 4.5 из 5 звезд4.5/5 (271)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersОт EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersРейтинг: 4.5 из 5 звезд4.5/5 (344)
- Team of Rivals: The Political Genius of Abraham LincolnОт EverandTeam of Rivals: The Political Genius of Abraham LincolnРейтинг: 4.5 из 5 звезд4.5/5 (234)
- The Unwinding: An Inner History of the New AmericaОт EverandThe Unwinding: An Inner History of the New AmericaРейтинг: 4 из 5 звезд4/5 (45)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyОт EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyРейтинг: 3.5 из 5 звезд3.5/5 (2219)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreОт EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreРейтинг: 4 из 5 звезд4/5 (1090)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)От EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Рейтинг: 4.5 из 5 звезд4.5/5 (119)
- S Y B Tech (Mechanical Engg) SyllabusДокумент30 страницS Y B Tech (Mechanical Engg) SyllabusBhavesh JainОценок пока нет
- Diversity of LifeДокумент6 страницDiversity of LifePriyanka VermaОценок пока нет
- Instant Download Ebook PDF An Introduction To Conservation Biology by Richard B Primack PDF ScribdДокумент41 страницаInstant Download Ebook PDF An Introduction To Conservation Biology by Richard B Primack PDF Scribdcharles.brookshire691100% (32)
- ICAIEB-2012 26th To 28th NovemberДокумент19 страницICAIEB-2012 26th To 28th NovemberChitrala KumaraswamynaiduОценок пока нет
- Goa University Syllabus - MSC - Zoology PDFДокумент43 страницыGoa University Syllabus - MSC - Zoology PDFdesaishantiОценок пока нет
- Areas System Act of 1992: National Integrated ProtectedДокумент51 страницаAreas System Act of 1992: National Integrated ProtectedRicardo Enriquez JrОценок пока нет
- Ecology TestДокумент17 страницEcology TestAbd Allah Emam Ewas EmamОценок пока нет
- Atalel GetuДокумент90 страницAtalel GetuBinyam BeyeneОценок пока нет
- Living Regents ReviewДокумент3 страницыLiving Regents Reviewsofia alamiaОценок пока нет
- Stutchbury - & - Morton - 2000 - Behavioral Ecology of Tropical Birds PDFДокумент169 страницStutchbury - & - Morton - 2000 - Behavioral Ecology of Tropical Birds PDFAnderson FelipeОценок пока нет
- Green Investments Required in The Forest Products Industry in NigeriaДокумент14 страницGreen Investments Required in The Forest Products Industry in NigeriaAlexander DeckerОценок пока нет
- Research Papers On Birds DiversityДокумент6 страницResearch Papers On Birds Diversityafnkjdhxlewftq100% (1)
- Lecture Note # 6 Biodiversity ConceptsДокумент17 страницLecture Note # 6 Biodiversity ConceptsBaucas, Rolanda D.Оценок пока нет
- Leu - The Myths of Safe Pesticides 2014Документ120 страницLeu - The Myths of Safe Pesticides 2014flexionОценок пока нет
- GEE 8 People and The Earths Ecosystem SyllabusДокумент7 страницGEE 8 People and The Earths Ecosystem SyllabusMarygin Esplana SarmientoОценок пока нет
- Service Functions of Agroforestry SystemsДокумент14 страницService Functions of Agroforestry SystemsJorge Rodríguez VásquezОценок пока нет
- Strengthening Climate Resilience in Siargao Islands Protected AreasДокумент164 страницыStrengthening Climate Resilience in Siargao Islands Protected AreasEpisode 17Оценок пока нет
- Major Threats to BiodiversityДокумент10 страницMajor Threats to BiodiversityCibi MОценок пока нет
- Importance of Industrial MineralsДокумент3 страницыImportance of Industrial MineralsElline Andreana ReyesОценок пока нет
- BS Botany 2012-13Документ78 страницBS Botany 2012-13Muhammad Rizwanullah TahirОценок пока нет
- 2020 Sustainable Agriculture Standard - Farm Requirements - Rainforest Alliance PDFДокумент83 страницы2020 Sustainable Agriculture Standard - Farm Requirements - Rainforest Alliance PDFLuis LanzasОценок пока нет
- Human-Induced Threats To Bio-Diversity in North-East India With Special Reference To MajuliДокумент10 страницHuman-Induced Threats To Bio-Diversity in North-East India With Special Reference To MajuliIJBSS,ISSN:2319-2968Оценок пока нет
- Executive Summary of Mt. Matutum Protected LandscapeДокумент5 страницExecutive Summary of Mt. Matutum Protected LandscapeDyan Geleen Ballesteros TubleОценок пока нет
- Biodiversity of Date Palm in The Sultanate of OmanДокумент7 страницBiodiversity of Date Palm in The Sultanate of OmanAl-Yahyai100% (14)
- International Rice Research Notes Vol.28 No.2Документ68 страницInternational Rice Research Notes Vol.28 No.2ccquintosОценок пока нет
- Diversity Management Critical Review and Future AgendaДокумент40 страницDiversity Management Critical Review and Future AgendaAnaОценок пока нет
- Evs ProjectДокумент19 страницEvs ProjectSaloni KariyaОценок пока нет
- Specimen Mark Scheme For Paper 1 For 2014 Onwards PDFДокумент6 страницSpecimen Mark Scheme For Paper 1 For 2014 Onwards PDFM. JosephОценок пока нет
- Đề Thi Chính Thức: Mục TiêuДокумент18 страницĐề Thi Chính Thức: Mục TiêuNgan GiangОценок пока нет
- The 7 Environmental PrinciplesДокумент11 страницThe 7 Environmental PrinciplesBeau MontОценок пока нет