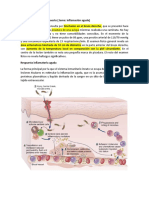
Вам также может понравиться
- Capitulo 11 InmunoДокумент26 страницCapitulo 11 InmunoJonatan PintoОценок пока нет
- Activacion Del Linfocito TДокумент8 страницActivacion Del Linfocito TMANUEL ALBERTO VERA PIMINCHUMOОценок пока нет
- Linfocitos T Cooperadores-1Документ5 страницLinfocitos T Cooperadores-1Jessica OrdoñezОценок пока нет
- Linfocito T CD4+ - Wikipedia, La Enciclopedia LibreДокумент13 страницLinfocito T CD4+ - Wikipedia, La Enciclopedia LibreShowerОценок пока нет
- Moléculas AccesoriasДокумент15 страницMoléculas AccesoriasPaola PintoОценок пока нет
- Resumen Cap 9 MurrayДокумент9 страницResumen Cap 9 MurrayMyle Campos LombeidaОценок пока нет
- Resumen Del Capítulo 9 de AbbasДокумент7 страницResumen Del Capítulo 9 de AbbasMaría Alejandra Peña100% (1)
- GUIA CLASE 12 2023-1 - Activacion Linfocitos TДокумент8 страницGUIA CLASE 12 2023-1 - Activacion Linfocitos TVale AlvarezОценок пока нет
- Linfocitos TДокумент15 страницLinfocitos TIGNACIO SALVADOR FERNANDEZ GIRONОценок пока нет
- Taller de InmunologíaДокумент5 страницTaller de InmunologíamayleОценок пока нет
- Clase 5Документ4 страницыClase 5Jesus Yepes martinezОценок пока нет
- Linfocitos TДокумент2 страницыLinfocitos TERIKA GISSELLE CUEVAS QUINTEROОценок пока нет
- Respuesta Funcional de Linfocito T Activados Fase de Contracciã N de La Respuesta de Cã© Lulas T SuperantigenosДокумент13 страницRespuesta Funcional de Linfocito T Activados Fase de Contracciã N de La Respuesta de Cã© Lulas T SuperantigenosPabsОценок пока нет
- Respuesta InmuneДокумент28 страницRespuesta Inmuneraziel boxОценок пока нет
- Linfocitos TДокумент4 страницыLinfocitos TPaulaAlvarezSuaОценок пока нет
- Resumen Inmunologia Introduccion A La Inmunologia VeterinariaДокумент12 страницResumen Inmunologia Introduccion A La Inmunologia VeterinariaAri MendozaОценок пока нет
- Respuestas Funcionales de Linfocitos T-Secreción de Citocinas y Expresión de Receptores para CitocinasДокумент6 страницRespuestas Funcionales de Linfocitos T-Secreción de Citocinas y Expresión de Receptores para CitocinasMelany GallardoОценок пока нет
- Tarea No.14 Proteínas Cd1 - Cd8 y TCR (Alfa y Beta) - Jorge Vargas Vargas Grupo 1 Tercer SemestreДокумент5 страницTarea No.14 Proteínas Cd1 - Cd8 y TCR (Alfa y Beta) - Jorge Vargas Vargas Grupo 1 Tercer SemestreJorge VargasОценок пока нет
- Maduración de Linfocitos TДокумент41 страницаMaduración de Linfocitos TVicToRia50% (2)
- Clase 8 Inmunidad Mediada Por CélulasДокумент42 страницыClase 8 Inmunidad Mediada Por CélulasYOCELINОценок пока нет
- Activación de Linfocitos TДокумент16 страницActivación de Linfocitos Tclarafra628Оценок пока нет
- Linfocito T VirgenДокумент3 страницыLinfocito T VirgensofiarodavilОценок пока нет
- Activacion Del Linfocitos TH y TCДокумент25 страницActivacion Del Linfocitos TH y TCJose Efraím Olvera LopezОценок пока нет
- Capítulo 13 Pavón. Linfocitos TДокумент12 страницCapítulo 13 Pavón. Linfocitos TClaudia PérezОценок пока нет
- PreguntasДокумент9 страницPreguntasSerCL1Оценок пока нет
- Linfopoyesis TimicaДокумент8 страницLinfopoyesis TimicaLilizhitaMaОценок пока нет
- Inmunidad 2Документ7 страницInmunidad 2joseОценок пока нет
- Primera Parte Taller de InmunologiaДокумент13 страницPrimera Parte Taller de Inmunologiajuan ariasОценок пока нет
- Celulas T HelperДокумент2 страницыCelulas T HelpermanualejandroОценок пока нет
- Activacion de Los Linfoncitos T.Документ6 страницActivacion de Los Linfoncitos T.Lis Juli MenReОценок пока нет
- Linfocitos T y Su Receptor (TCR)Документ8 страницLinfocitos T y Su Receptor (TCR)Giovanna B100% (11)
- C2 Inmunologia CompletoДокумент76 страницC2 Inmunologia CompletoDiego Alejandro Muñoz ChinchillaОценок пока нет
- C2 - Laboratorio de InmunologiaДокумент53 страницыC2 - Laboratorio de InmunologiaDiego Alejandro Muñoz ChinchillaОценок пока нет
- Respuesta Inmune AdaptativaДокумент11 страницRespuesta Inmune Adaptativaalessandra.sandoval.93Оценок пока нет
- Maduración y Activación de Linfocitos TДокумент59 страницMaduración y Activación de Linfocitos TVioleta Tello MedinaОценок пока нет
- Linfocito T y CitocinasДокумент28 страницLinfocito T y CitocinasAlfredo VidalesОценок пока нет
- Revisión BibliográficaДокумент9 страницRevisión BibliográficasofiaОценок пока нет
- Resumen Cap 6 Inmunologia de AbbasДокумент7 страницResumen Cap 6 Inmunologia de Abbasmontsesitacm1104Оценок пока нет
- Activación de Linfocitos TДокумент26 страницActivación de Linfocitos TFernandho Cruz BaciiliioОценок пока нет
- Capitulo 12 InmunologiaДокумент22 страницыCapitulo 12 InmunologiaCarLiita CruzОценок пока нет
- Resumen-Activación Del Linfocito T (Corregido)Документ4 страницыResumen-Activación Del Linfocito T (Corregido)alisson louridoОценок пока нет
- Celulas y Tejidos Sel Sistema RioДокумент42 страницыCelulas y Tejidos Sel Sistema Riochivas22Оценок пока нет
- Linfocitos CD4 y CD8Документ4 страницыLinfocitos CD4 y CD8Victor Eduardo Jimenez ValdezОценок пока нет
- Citotoxicidad PDFДокумент2 страницыCitotoxicidad PDFOctavio ZmОценок пока нет
- Linfocito TДокумент56 страницLinfocito TRonyzhito CondoriОценок пока нет
- Generación de Linfocitos T EfectoresДокумент11 страницGeneración de Linfocitos T EfectoresMariano Félix DemasiОценок пока нет
- Casos Clínicos SI 202310Документ46 страницCasos Clínicos SI 202310Avery DreamsОценок пока нет
- Resumen Cap 6 InmunoДокумент6 страницResumen Cap 6 Inmunobopeepout papuОценок пока нет
- Cap. 5 - Inmunidad Mediada Por Linfocitos TДокумент30 страницCap. 5 - Inmunidad Mediada Por Linfocitos TDaviid A'SОценок пока нет
- Celulas THДокумент5 страницCelulas THmonse_moonОценок пока нет
- Clase 3 Linfocito B y T SEMANA 3Документ64 страницыClase 3 Linfocito B y T SEMANA 3OCTAVIOОценок пока нет
- Inmunologia Linfocitos T 13 Sep 2021Документ5 страницInmunologia Linfocitos T 13 Sep 2021Victor Eduardo Jimenez ValdezОценок пока нет
- Celulas TДокумент12 страницCelulas TStephany Guadalupe Soto GarciaОценок пока нет
- Inmuno V Colaboracin T-BДокумент5 страницInmuno V Colaboracin T-BLuna LorenzoОценок пока нет
- Clase 4 Linfocitos T 2Документ13 страницClase 4 Linfocitos T 2fialophora0210Оценок пока нет
- Activación de LinfocitosДокумент25 страницActivación de Linfocitoselizabeth890716Оценок пока нет
- Exposicion de Micro II de Funcion de Linfocitos T y BДокумент25 страницExposicion de Micro II de Funcion de Linfocitos T y Bjubachar3128Оценок пока нет
- MORFO2Документ8 страницMORFO2ValeriaNadjarОценок пока нет
- Ventilacion - Neonatal IMPORTANTISIMO PDFДокумент94 страницыVentilacion - Neonatal IMPORTANTISIMO PDFJuan Carlos HuanacheaОценок пока нет
- Bipedestador Paralelogramo PDFДокумент10 страницBipedestador Paralelogramo PDFAxel RamosОценок пока нет
- Tubo DigestivoДокумент1 страницаTubo DigestivoCarolina VillarroelОценок пока нет
- Ingredientes de Suplementos Dietéticos Con Efectos Benéficos para La Salud Clínicamente DocumentadosДокумент274 страницыIngredientes de Suplementos Dietéticos Con Efectos Benéficos para La Salud Clínicamente DocumentadosCarlos Eduardo Castañeda Muñoz100% (1)
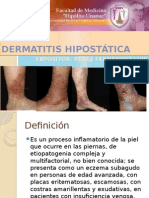
- Dermatitis HipostáticaДокумент14 страницDermatitis HipostáticaLuis Angel Perez FernandezОценок пока нет
- Plan Clase No 1 - Introducción A La Tecnología Farmacéutica y Estudios de Pre Formulación 8 A 12-10-18 (1) PDFДокумент42 страницыPlan Clase No 1 - Introducción A La Tecnología Farmacéutica y Estudios de Pre Formulación 8 A 12-10-18 (1) PDFNozomi MiloОценок пока нет
- 1806 5887 2 PBДокумент6 страниц1806 5887 2 PBJhonn RamosОценок пока нет
- Taller de BiologiaДокумент25 страницTaller de BiologiaÄŃĐËŘŠÖŇ ÅŘĆŐŚОценок пока нет
- Grupo N°2 - Diagrama de FlujoДокумент4 страницыGrupo N°2 - Diagrama de FlujoMaria Emilia MedinaОценок пока нет
- Principios Del Metabolismo MicrobianoДокумент32 страницыPrincipios Del Metabolismo MicrobianoKarina Vargas100% (1)
- Disección de DorsoДокумент3 страницыDisección de Dorsomiguelperedogonzalez41099Оценок пока нет
- 160 - Rabdiomiolisis Como Complicacion en Las Urgencias Por Quemaduras Electricas. Presentacion de Un Caso PDFДокумент12 страниц160 - Rabdiomiolisis Como Complicacion en Las Urgencias Por Quemaduras Electricas. Presentacion de Un Caso PDFRaul CerratoОценок пока нет
- Mapa Metabólico UanДокумент1 страницаMapa Metabólico UanJONATHAN SAUL HERRERA RAMIREZ100% (1)
- Estimulación TempranaДокумент60 страницEstimulación Tempranajanet_kp100% (3)
- Cuestionario #16 de BioquímicaДокумент2 страницыCuestionario #16 de BioquímicaYazminPotterОценок пока нет
- Prueba de Ciencia Naturales Aparato ExcretorДокумент6 страницPrueba de Ciencia Naturales Aparato ExcretorarturcossОценок пока нет
- Sistema Nervioso WordДокумент9 страницSistema Nervioso WordKevin AguirreОценок пока нет
- Ruiz Torres Raul PavelДокумент102 страницыRuiz Torres Raul PavelJhoNes IbarraОценок пока нет
- UrinarioДокумент5 страницUrinario22954Оценок пока нет
- Fisio ExposicionДокумент7 страницFisio ExposicionJose Antonio Solis ChipaОценок пока нет
- Estructura Externa de La BacteriaДокумент10 страницEstructura Externa de La BacteriaHoss SeeeОценок пока нет
- 01MTEG BSerrano PDFДокумент131 страница01MTEG BSerrano PDFNo tengo nombre100% (1)
- TP4 - Hombro y Sus Articulaciones - Ud2Документ4 страницыTP4 - Hombro y Sus Articulaciones - Ud2Edson PereiraОценок пока нет
- 4to Informe (Visceras de Pollo)Документ6 страниц4to Informe (Visceras de Pollo)Eduardo Antonio Molinari NovoaОценок пока нет
- Grant Anatomia para ColorearДокумент560 страницGrant Anatomia para ColorearLITZE ZULEMA CHURATA LUNAОценок пока нет
- Informe, Recuento LeucocitarioДокумент15 страницInforme, Recuento LeucocitarioStephanieОценок пока нет
- OsteologiaДокумент24 страницыOsteologiaValdiviezo MujicaОценок пока нет
- Concepto y Fisiopatologia de La NeumoniaДокумент6 страницConcepto y Fisiopatologia de La NeumoniaAriana BendezúОценок пока нет
- 09 PDFДокумент17 страниц09 PDFlicherjdk100% (1)
- Clase 1Документ15 страницClase 1CarlonchaCáceresОценок пока нет