Вам также может понравиться
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryОт EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryРейтинг: 3.5 из 5 звезд3.5/5 (231)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)От EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Рейтинг: 4.5 из 5 звезд4.5/5 (119)
- Never Split the Difference: Negotiating As If Your Life Depended On ItОт EverandNever Split the Difference: Negotiating As If Your Life Depended On ItРейтинг: 4.5 из 5 звезд4.5/5 (838)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaОт EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaРейтинг: 4.5 из 5 звезд4.5/5 (265)
- The Little Book of Hygge: Danish Secrets to Happy LivingОт EverandThe Little Book of Hygge: Danish Secrets to Happy LivingРейтинг: 3.5 из 5 звезд3.5/5 (399)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyОт EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyРейтинг: 3.5 из 5 звезд3.5/5 (2219)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeОт EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeРейтинг: 4 из 5 звезд4/5 (5794)
- Team of Rivals: The Political Genius of Abraham LincolnОт EverandTeam of Rivals: The Political Genius of Abraham LincolnРейтинг: 4.5 из 5 звезд4.5/5 (234)
- The Emperor of All Maladies: A Biography of CancerОт EverandThe Emperor of All Maladies: A Biography of CancerРейтинг: 4.5 из 5 звезд4.5/5 (271)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreОт EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreРейтинг: 4 из 5 звезд4/5 (1090)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersОт EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersРейтинг: 4.5 из 5 звезд4.5/5 (344)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceОт EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceРейтинг: 4 из 5 звезд4/5 (894)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureОт EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureРейтинг: 4.5 из 5 звезд4.5/5 (474)
- Easy Gluten Free RecipesДокумент90 страницEasy Gluten Free RecipesBrandon Schmid100% (1)
- The Unwinding: An Inner History of the New AmericaОт EverandThe Unwinding: An Inner History of the New AmericaРейтинг: 4 из 5 звезд4/5 (45)
- The Yellow House: A Memoir (2019 National Book Award Winner)От EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Рейтинг: 4 из 5 звезд4/5 (98)
- A Comparative Look at Jamaican Creole and Guyanese CreoleДокумент18 страницA Comparative Look at Jamaican Creole and Guyanese CreoleShivana Allen100% (3)
- Financial Management-Capital BudgetingДокумент39 страницFinancial Management-Capital BudgetingParamjit Sharma100% (53)
- CRUSADE of PRAYERS 1-170 Litany 1-6 For The Key To Paradise For DistributionДокумент264 страницыCRUSADE of PRAYERS 1-170 Litany 1-6 For The Key To Paradise For DistributionJESUS IS RETURNING DURING OUR GENERATION100% (10)
- Week 4 CasesДокумент181 страницаWeek 4 CasesMary Ann AmbitaОценок пока нет
- Strategies in Various Speech SituationsДокумент6 страницStrategies in Various Speech SituationsSky NayytОценок пока нет
- BioremediasiДокумент9 страницBioremediasifireblack37Оценок пока нет
- PNAS 1992 Frosch 800 3Документ5 страницPNAS 1992 Frosch 800 3fireblack37Оценок пока нет
- Biosurfactant-Enhanced Hydrocarbon Bioremediation An OverviewДокумент7 страницBiosurfactant-Enhanced Hydrocarbon Bioremediation An Overviewfireblack37Оценок пока нет
- 2014 - Bioremediation of Petroleum Hydrocarbons Catabolic Genes, Microbial Communities, and ApplicationsДокумент14 страниц2014 - Bioremediation of Petroleum Hydrocarbons Catabolic Genes, Microbial Communities, and Applicationsfireblack37Оценок пока нет
- Ijms 12 04550Документ24 страницыIjms 12 04550Dr-AdnanAMalikОценок пока нет
- A Mechanistic Overview of Health Associated Effects of Low Levels of Organochlorine and Organophosphorous Pesticides.Документ6 страницA Mechanistic Overview of Health Associated Effects of Low Levels of Organochlorine and Organophosphorous Pesticides.fireblack37Оценок пока нет
- Acetylcholinesterase Inhibitors Pharmacology and ToxicologyДокумент21 страницаAcetylcholinesterase Inhibitors Pharmacology and Toxicologyfireblack37Оценок пока нет
- Sensor and ActuatorsДокумент8 страницSensor and Actuatorsfireblack37Оценок пока нет
- Ijms 12 04550Документ24 страницыIjms 12 04550Dr-AdnanAMalikОценок пока нет
- 2014, Mavunganidze, Madakadze, Nyamangara and Mafongoya, Impact of Tillage On Weeds, Crop Protection 1-s2 0-S0261219413003487-MainДокумент8 страниц2014, Mavunganidze, Madakadze, Nyamangara and Mafongoya, Impact of Tillage On Weeds, Crop Protection 1-s2 0-S0261219413003487-Mainfireblack37Оценок пока нет
- Ijms 12 04550Документ24 страницыIjms 12 04550Dr-AdnanAMalikОценок пока нет
- Jurnal Utama FishewДокумент16 страницJurnal Utama Fishewfireblack37Оценок пока нет
- 3 AsdasdsaДокумент5 страниц3 Asdasdsafireblack37Оценок пока нет
- Variasi Intraspesifik Dalam Kecepatan Tumbuh Di Antara Tiga Populasi Gastropoda Intertidal, Nerita Japonica (Dunker)Документ9 страницVariasi Intraspesifik Dalam Kecepatan Tumbuh Di Antara Tiga Populasi Gastropoda Intertidal, Nerita Japonica (Dunker)Prince LeivyОценок пока нет
- Explore the rules and history of American football and basketballДокумент2 страницыExplore the rules and history of American football and basketballAndrei IoneanuОценок пока нет
- Spain Usa ExtraditionДокумент2 страницыSpain Usa ExtraditionAdrian BirdeaОценок пока нет
- Krittika Takiar PDFДокумент2 страницыKrittika Takiar PDFSudhakar TomarОценок пока нет
- Chapter 6 Sequence PakistanДокумент16 страницChapter 6 Sequence PakistanAsif Ullah0% (1)
- Land Securities Group (A)Документ13 страницLand Securities Group (A)Piyush SamalОценок пока нет
- All EscortsДокумент8 страницAll Escortsvicky19937Оценок пока нет
- Sosai Masutatsu Oyama - Founder of Kyokushin KarateДокумент9 страницSosai Masutatsu Oyama - Founder of Kyokushin KarateAdriano HernandezОценок пока нет
- Anschutz Nautopilot 5000Документ4 страницыAnschutz Nautopilot 5000Văn Phú PhạmОценок пока нет
- How To Write A Cover Letter For A Training ProgramДокумент4 страницыHow To Write A Cover Letter For A Training Programgyv0vipinem3100% (2)
- A Model For Blockchain-Based Distributed Electronic Health Records - 2016Документ14 страницA Model For Blockchain-Based Distributed Electronic Health Records - 2016Asif KhalidОценок пока нет
- Final Revised ResearchДокумент35 страницFinal Revised ResearchRia Joy SanchezОценок пока нет
- StoreFront 3.11Документ162 страницыStoreFront 3.11AnonimovОценок пока нет
- The Man of Sorrows Wednesday of Holy Week Divine IntimacyДокумент5 страницThe Man of Sorrows Wednesday of Holy Week Divine IntimacyTerri ThomasОценок пока нет
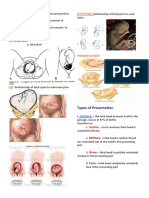
- The 4Ps of Labor: Passenger, Passageway, Powers, and PlacentaДокумент4 страницыThe 4Ps of Labor: Passenger, Passageway, Powers, and PlacentaMENDIETA, JACQUELINE V.Оценок пока нет
- DC Rebirth Reading Order 20180704Документ43 страницыDC Rebirth Reading Order 20180704Michael MillerОценок пока нет
- Understanding Malaysian Property TaxationДокумент68 страницUnderstanding Malaysian Property TaxationLee Chee KheongОценок пока нет
- Dell Market ResearchДокумент27 страницDell Market ResearchNaved Deshmukh0% (1)
- Zeal Study 10th English Synonym Unit 1 - 7Документ24 страницыZeal Study 10th English Synonym Unit 1 - 7viaanenterprises2008Оценок пока нет
- AIESEC - Exchange Participant (EP) GuidebookДокумент24 страницыAIESEC - Exchange Participant (EP) GuidebookAnonymous aoQ8gc1Оценок пока нет
- Aspartame Literature ReviewДокумент10 страницAspartame Literature Reviewapi-272556824Оценок пока нет
- 6-Modul - PTI412 - IoT-1-2020Документ17 страниц6-Modul - PTI412 - IoT-1-202020200801015 Joko SulistyoОценок пока нет
- Unit 2 - BT MLH 11 - KeyДокумент2 страницыUnit 2 - BT MLH 11 - KeyttyannieОценок пока нет
- Chapter 3 - The Role of Multimedia in EducationДокумент37 страницChapter 3 - The Role of Multimedia in EducationGellie Ann Silvestre RapinОценок пока нет
- Ujian NasionalДокумент41 страницаUjian NasionalKeisha SalsabilaОценок пока нет