Вам также может понравиться
- Pericyte Biology - Novel ConceptsОт EverandPericyte Biology - Novel ConceptsAlexander BirbrairОценок пока нет
- Brief Definitive ReportДокумент6 страницBrief Definitive ReportenilОценок пока нет
- Identification and Characterization of Circulating Human Transitional B CellsДокумент9 страницIdentification and Characterization of Circulating Human Transitional B CellsLizeth Rincon DelgadoОценок пока нет
- And Preferential Recruitment of B1 Cells in The Defective B1 Cell Homing To The Peritoneal CavityДокумент8 страницAnd Preferential Recruitment of B1 Cells in The Defective B1 Cell Homing To The Peritoneal Cavityflaviu_tabaranОценок пока нет
- A Comparison of Lymphocyte Micro Nuclei and Plasma Micro Nutrients in Vegetarians and Non-VegetariansДокумент8 страницA Comparison of Lymphocyte Micro Nuclei and Plasma Micro Nutrients in Vegetarians and Non-VegetariansNarcissus CassanovaОценок пока нет
- Cells 08 00724Документ13 страницCells 08 00724meltwithsnow163.comОценок пока нет
- Ni Hms 52672Документ18 страницNi Hms 52672Patricia BenevidesОценок пока нет
- Braniste 2014Документ24 страницыBraniste 20142021207549Оценок пока нет
- 17d Stem Cells ScieMedPoliticalДокумент4 страницы17d Stem Cells ScieMedPoliticalbbdddОценок пока нет
- Biology of Blood and Marrow Transplantation: ReviewsДокумент19 страницBiology of Blood and Marrow Transplantation: Reviewsmuhammad fadliОценок пока нет
- Fragi 02 797040Документ16 страницFragi 02 797040guischmoОценок пока нет
- Reproductive Systems Biology Tackles Global Issues of Population Growth, Food Safety and Reproductive HealthДокумент5 страницReproductive Systems Biology Tackles Global Issues of Population Growth, Food Safety and Reproductive HealthElla B. CollantesОценок пока нет
- Medical InfosДокумент9 страницMedical InfosPaul AsturbiarisОценок пока нет
- Long Term Engraftment of ESC Derived B 1 Progenitor Cells Su 2019 Stem CellДокумент12 страницLong Term Engraftment of ESC Derived B 1 Progenitor Cells Su 2019 Stem CellLuisa FernandaОценок пока нет
- A Switch in Time Through Genes Aligned Unraveling The Geno - 2020 - Cell StemДокумент3 страницыA Switch in Time Through Genes Aligned Unraveling The Geno - 2020 - Cell StemBandar Ternama XBОценок пока нет
- Dev Bio IntroductoryДокумент17 страницDev Bio Introductoryreynaldo banaria jrОценок пока нет
- Blood JournalДокумент12 страницBlood Journaltammy_tataОценок пока нет
- Maduracion Linfocito BДокумент12 страницMaduracion Linfocito BAngie Regalado SenmacheОценок пока нет
- PIIS2213671114002951Документ15 страницPIIS2213671114002951Ana LimeiraОценок пока нет
- JEM InnateImmunity Special IssueДокумент92 страницыJEM InnateImmunity Special IssueAnonymous 6OPLC9UОценок пока нет
- Phil. Trans. R. Soc. B 2011 Unternaehrer 2274 85Документ13 страницPhil. Trans. R. Soc. B 2011 Unternaehrer 2274 85Jo-Anne M. VentiganОценок пока нет
- Myeloid Cell Origins, Differentiation, and Clinical ImplicationsДокумент18 страницMyeloid Cell Origins, Differentiation, and Clinical ImplicationsWendy PОценок пока нет
- Ncomms12015 Scleroza Multipla PDFДокумент11 страницNcomms12015 Scleroza Multipla PDFLUTICHIEVICI NATALIAОценок пока нет
- Biofilms and Chronic Infections: CommentariesДокумент3 страницыBiofilms and Chronic Infections: CommentariesnissaОценок пока нет
- SickKids Research ProgramsДокумент44 страницыSickKids Research ProgramsKlutz4LifeОценок пока нет
- 1 s2.0 S0012160620303195 Main 1 PDFДокумент20 страниц1 s2.0 S0012160620303195 Main 1 PDFNabila NajwaОценок пока нет
- Berberine Induces Caspase-Independent Cell Death in Colon Tumor Cells Through Activation of Apoptosis-Inducing FactorДокумент12 страницBerberine Induces Caspase-Independent Cell Death in Colon Tumor Cells Through Activation of Apoptosis-Inducing FactorflaiharaОценок пока нет
- 1471 2431 11 29 PDFДокумент6 страниц1471 2431 11 29 PDFCeleste SantiОценок пока нет
- Advances in Reprogramming Somatic Cells To InducedДокумент21 страницаAdvances in Reprogramming Somatic Cells To InducedNatalia MoysésОценок пока нет
- 2020 Frontiers Cell Dev Biol Soares-da-Silva F Etal Al.Документ20 страниц2020 Frontiers Cell Dev Biol Soares-da-Silva F Etal Al.PerpetuaОценок пока нет
- Atlas RNAДокумент42 страницыAtlas RNArenyuandshisenyuanОценок пока нет
- Clonacion ...Документ9 страницClonacion ...kirly_19Оценок пока нет
- Sex Is Binary PIP 12 2020Документ2 страницыSex Is Binary PIP 12 2020Hayden LukasОценок пока нет
- Bone Marrow Evaluation For Pediatric PatientsДокумент7 страницBone Marrow Evaluation For Pediatric PatientssylvetriОценок пока нет
- Author's Accepted Manuscript: Seminars in Pediatric SurgeryДокумент28 страницAuthor's Accepted Manuscript: Seminars in Pediatric Surgerywibm23Оценок пока нет
- Embryonic Stem Cells ThesisДокумент6 страницEmbryonic Stem Cells Thesisqpftgehig100% (2)
- Nature 11647Документ5 страницNature 11647Jef_8Оценок пока нет
- And + Peptide That Is Recognized by Human CD4 Characterization of AДокумент13 страницAnd + Peptide That Is Recognized by Human CD4 Characterization of ABiosynthesisОценок пока нет
- B Cells in Immunity and Tolerance - BookДокумент190 страницB Cells in Immunity and Tolerance - BookJuan FranciscoОценок пока нет
- 2015 Proceedings of The Second Meeting of The French-Speaking Society SF-DOHaDДокумент50 страниц2015 Proceedings of The Second Meeting of The French-Speaking Society SF-DOHaDNguyễn Tiến HồngОценок пока нет
- Classifying HumansДокумент2 страницыClassifying HumansAnonymous uliBzsZrWОценок пока нет
- Wharton's JellyДокумент13 страницWharton's JellyZ MОценок пока нет
- Cytometry Part B Clinical - 2018 - Garcia Prat - Extended Immunophenotyping Reference Values in A Healthy PediatricДокумент11 страницCytometry Part B Clinical - 2018 - Garcia Prat - Extended Immunophenotyping Reference Values in A Healthy PediatricSabina-Gabriela MihaiОценок пока нет
- @medicinejournal American Journal of Perinatology September 2017Документ128 страниц@medicinejournal American Journal of Perinatology September 2017angsukriОценок пока нет
- Red Biotechnology: A Healthy World: Elsayed I.G., Kanwugu O.N., Ivantsova M.NДокумент4 страницыRed Biotechnology: A Healthy World: Elsayed I.G., Kanwugu O.N., Ivantsova M.NWrexford BritneyОценок пока нет
- The Vaginal Microbiome New Information About Genital Tract Flora Using Molecular Based TechniquesДокумент17 страницThe Vaginal Microbiome New Information About Genital Tract Flora Using Molecular Based TechniquesJing XueОценок пока нет
- Apoptosis and Expression of Bcl-2 and Bax During Early Human EmbryogenesisДокумент6 страницApoptosis and Expression of Bcl-2 and Bax During Early Human EmbryogenesisCita BudiartiОценок пока нет
- Research Paper Embryonic Stem CellsДокумент8 страницResearch Paper Embryonic Stem Cellsrrndfrrif100% (1)
- Evidence Supporting The Biological Nature of Gender IdentityДокумент6 страницEvidence Supporting The Biological Nature of Gender IdentityMorgan Atwood100% (2)
- Evidence For Two Types of Brown Adipose Tissue in Humans: LettersДокумент5 страницEvidence For Two Types of Brown Adipose Tissue in Humans: LettersgotokoreaОценок пока нет
- Peopling The AmericasДокумент3 страницыPeopling The Americasmikey_tipswordОценок пока нет
- Mouse ModelsДокумент32 страницыMouse ModelsCiocan AlexandraОценок пока нет
- Ismej 2008 108Документ11 страницIsmej 2008 108Alex Cristian IonutОценок пока нет
- Hassan 2008Документ4 страницыHassan 2008Sabrina ParenteОценок пока нет
- Ectopic Lymphoid Follicle Formation and Human Seasonal Influenza Vaccination ResponsesДокумент15 страницEctopic Lymphoid Follicle Formation and Human Seasonal Influenza Vaccination Responseslimengdi0269Оценок пока нет
- Undulating Changes in Human Plasma Proteome Profiles Across The LifespanДокумент22 страницыUndulating Changes in Human Plasma Proteome Profiles Across The LifespanЗлойБобрОценок пока нет
- Biomarkers Pediatric TBДокумент9 страницBiomarkers Pediatric TBPaulo MattosОценок пока нет
- Aggressive Lymphomas NEJMДокумент13 страницAggressive Lymphomas NEJMAnnaОценок пока нет
- Leydig Cells TrasplantДокумент28 страницLeydig Cells TrasplantKevin VillaОценок пока нет
- Architecture of The Outbred Brown Fat Proteome DefДокумент49 страницArchitecture of The Outbred Brown Fat Proteome Defh5wwgbvbvmОценок пока нет
- Gromacs Tutorial For Drug Enzyme ComplexДокумент21 страницаGromacs Tutorial For Drug Enzyme ComplexManoj Kumar RoutОценок пока нет
- Gromacs Tutorial For Drug Enzyme ComplexДокумент21 страницаGromacs Tutorial For Drug Enzyme ComplexManoj Kumar RoutОценок пока нет
- Gromacs Tutorial For Drug Enzyme ComplexДокумент21 страницаGromacs Tutorial For Drug Enzyme ComplexManoj Kumar RoutОценок пока нет
- Gromacs Tutorial For Drug Enzyme ComplexДокумент21 страницаGromacs Tutorial For Drug Enzyme ComplexManoj Kumar RoutОценок пока нет
- Drugs and Intellectual Property Right1Документ3 страницыDrugs and Intellectual Property Right1rajagopalan86Оценок пока нет
- Dosage Lec - Unit 2 Drug DiscoveryДокумент8 страницDosage Lec - Unit 2 Drug DiscoveryJohn Lorenz BordadorОценок пока нет
- Theories, Models, and FrameworksДокумент24 страницыTheories, Models, and FrameworksLemuel LinoОценок пока нет
- Macro Calorie Calculator TemplateДокумент23 страницыMacro Calorie Calculator TemplateCarla BaraybarОценок пока нет
- Quality Laboratory Standards: in KenyaДокумент20 страницQuality Laboratory Standards: in KenyaKate NjinuОценок пока нет
- Neurotology Fellowship MilestonesДокумент18 страницNeurotology Fellowship MilestonesAnonymous AuJncFVWvОценок пока нет
- Understanding Receiver Operating Characteristic Roc CurvesДокумент2 страницыUnderstanding Receiver Operating Characteristic Roc CurvesOlievia Herinda AdhiafebriantyОценок пока нет
- Female PelvisДокумент11 страницFemale PelvisAhmed Gh Al-zechrawi100% (1)
- NCP Self HarmДокумент1 страницаNCP Self HarmJoshua ArevaloОценок пока нет
- Research Paper Final DraftДокумент17 страницResearch Paper Final Draftapi-740628337Оценок пока нет
- 10 Critical Care PDF FreeДокумент3 страницы10 Critical Care PDF FreeAdnene TouatiОценок пока нет
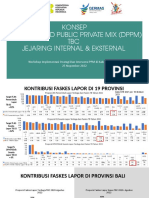
- Konsep District Based Public Private Mix (DPPM) TBC Jejaring Internal & EksternalДокумент24 страницыKonsep District Based Public Private Mix (DPPM) TBC Jejaring Internal & EksternalMade SupadmaОценок пока нет
- 2020 Drug PricesДокумент25 страниц2020 Drug PricesChris-Goldie LorezoОценок пока нет
- Jet Lag PDFДокумент8 страницJet Lag PDFIan FitzpatrickОценок пока нет
- 2010 Acr Criteria PDFДокумент3 страницы2010 Acr Criteria PDFMuhammad Abdul RahmanОценок пока нет
- Himalaya PublicationДокумент20 страницHimalaya Publicationanand1540Оценок пока нет
- Tanganyika Nacte Past Paper-1Документ116 страницTanganyika Nacte Past Paper-1Mnawaru KhalfaniОценок пока нет
- 7 Stars DoctorДокумент2 страницы7 Stars DoctorCynthia Yupa100% (1)
- Reality TherapyДокумент1 страницаReality TherapyTinoRepaso100% (1)
- FullmanuscriptДокумент3 страницыFullmanuscriptAmalia Tri UtamiОценок пока нет
- Distal ExtensionДокумент110 страницDistal ExtensionParmod GuliaОценок пока нет
- Chikungunya IgM Combo Rev DДокумент2 страницыChikungunya IgM Combo Rev DDaniel LaraОценок пока нет
- Medical Ethics More Than Hundred CasesДокумент33 страницыMedical Ethics More Than Hundred Caseskrebkreb100% (4)
- Drug-Induced Nutrient DepletionДокумент4 страницыDrug-Induced Nutrient DepletionVictoria HristovaОценок пока нет
- Precedex DosingДокумент4 страницыPrecedex DosingnonemiscОценок пока нет
- PTIДокумент19 страницPTIDick Jerzy Salazar FranciaОценок пока нет
- Trisomy 21Документ5 страницTrisomy 21pongidaeОценок пока нет
- Beneficence and Non-MaleficenceДокумент3 страницыBeneficence and Non-MaleficenceRathin BanerjeeОценок пока нет
- 2Документ78 страниц2api-3744136Оценок пока нет
- Msds Asam MefenamatДокумент6 страницMsds Asam MefenamatKristal Resa Diena RОценок пока нет
- Next To Normal - Abridged ScriptДокумент13 страницNext To Normal - Abridged Scriptterrify007Оценок пока нет