Вам также может понравиться
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeОт EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeРейтинг: 4 из 5 звезд4/5 (5794)
- The Little Book of Hygge: Danish Secrets to Happy LivingОт EverandThe Little Book of Hygge: Danish Secrets to Happy LivingРейтинг: 3.5 из 5 звезд3.5/5 (399)
- Troncoso-PalaciosETAL2018-Phymaturus Loboi Sp. Nov.Документ20 страницTroncoso-PalaciosETAL2018-Phymaturus Loboi Sp. Nov.laspiur22blues7327Оценок пока нет
- Troncoso PalaciosETAL2013 Phymaturus BibroniiДокумент15 страницTroncoso PalaciosETAL2013 Phymaturus Bibroniilaspiur22blues7327Оценок пока нет
- Sanabria, Quiroga & Laspiur-ODONTOPHRYNUS OCCIDENTALIS-Scoliosis and Partial Albinism-2010Документ2 страницыSanabria, Quiroga & Laspiur-ODONTOPHRYNUS OCCIDENTALIS-Scoliosis and Partial Albinism-2010laspiur22blues7327Оценок пока нет
- Laspiur & Acosta-LIOLAMUS CUYANUS-Clutch Size 2008Документ2 страницыLaspiur & Acosta-LIOLAMUS CUYANUS-Clutch Size 2008laspiur22blues7327Оценок пока нет
- Snake Copulation Observation Provides Insights into Reproductive Behavior of Threatened West Indian SpeciesДокумент2 страницыSnake Copulation Observation Provides Insights into Reproductive Behavior of Threatened West Indian Specieslaspiur22blues7327Оценок пока нет
- NATURAL PREDATORДокумент2 страницыNATURAL PREDATORlaspiur22blues7327Оценок пока нет
- Cabezas Cartes Et Al-Reproduction Phymaturus-2010Документ11 страницCabezas Cartes Et Al-Reproduction Phymaturus-2010laspiur22blues7327Оценок пока нет
- Sanabria, Quiroga & Laspiur-ODONTOPHRYNUS OCCIDENTALIS-Scoliosis and Partial Albinism-2010Документ2 страницыSanabria, Quiroga & Laspiur-ODONTOPHRYNUS OCCIDENTALIS-Scoliosis and Partial Albinism-2010laspiur22blues7327Оценок пока нет
- Laspiur & Acosta 2015-Leiosauus Jaguaris Distribution PDFДокумент4 страницыLaspiur & Acosta 2015-Leiosauus Jaguaris Distribution PDFlaspiur22blues7327Оценок пока нет
- AbdalaETAL2017 Liolaemus Rabinoi PDFДокумент12 страницAbdalaETAL2017 Liolaemus Rabinoi PDFlaspiur22blues7327Оценок пока нет
- Troncoso PalaciosETAL2013 Phymaturus BibroniiДокумент15 страницTroncoso PalaciosETAL2013 Phymaturus Bibroniilaspiur22blues7327Оценок пока нет
- LaspiurETal2010 Boa PDFДокумент2 страницыLaspiurETal2010 Boa PDFlaspiur22blues7327Оценок пока нет
- Kohl Et Al-2016-Molecular EcologyДокумент15 страницKohl Et Al-2016-Molecular Ecologylaspiur22blues7327Оценок пока нет
- Gómez AlesETAL2017Документ10 страницGómez AlesETAL2017laspiur22blues7327Оценок пока нет
- LaspiurETal2010 OdontophrynusДокумент1 страницаLaspiurETal2010 Odontophrynuslaspiur22blues7327Оценок пока нет
- LaspiurETal2010 OdontophrynusДокумент1 страницаLaspiurETal2010 Odontophrynuslaspiur22blues7327Оценок пока нет
- Snake Copulation Observation Provides Insights into Reproductive Behavior of Threatened West Indian SpeciesДокумент2 страницыSnake Copulation Observation Provides Insights into Reproductive Behavior of Threatened West Indian Specieslaspiur22blues7327Оценок пока нет
- Laspiur, Villavicencio & Acosta-LIOLAEMUS CHACOENSIS-Body Temperature-2008Документ2 страницыLaspiur, Villavicencio & Acosta-LIOLAEMUS CHACOENSIS-Body Temperature-2008laspiur22blues7327Оценок пока нет
- Snake Copulation Observation Provides Insights into Reproductive Behavior of Threatened West Indian SpeciesДокумент2 страницыSnake Copulation Observation Provides Insights into Reproductive Behavior of Threatened West Indian Specieslaspiur22blues7327Оценок пока нет
- Gómez Et Al 2013Документ1 страницаGómez Et Al 2013laspiur22blues7327Оценок пока нет
- Caicedo-Portilla Et Al-2010-Diet, Microhabitat Use Daily Activity Patterns of An Andean Population of Mabuya (Squamata, Scincidae) - SAJHДокумент7 страницCaicedo-Portilla Et Al-2010-Diet, Microhabitat Use Daily Activity Patterns of An Andean Population of Mabuya (Squamata, Scincidae) - SAJHlaspiur22blues7327Оценок пока нет
- Laspiur & Acosta 2015-Leiosaurus Jaguaris DistributionДокумент4 страницыLaspiur & Acosta 2015-Leiosaurus Jaguaris Distributionlaspiur22blues7327Оценок пока нет
- Bouin'S Fixative (Bouin'S Fluid) : Technical Data Sheet 866Документ1 страницаBouin'S Fixative (Bouin'S Fluid) : Technical Data Sheet 866laspiur22blues7327Оценок пока нет
- EvolDevelop 2013 15 p326Документ18 страницEvolDevelop 2013 15 p326laspiur22blues7327Оценок пока нет
- Herzog Et Al 2003. Seasonal Variation in Avian Community Composition in A High-Andean Polylepis (Rosaceae) Forest FragmentДокумент10 страницHerzog Et Al 2003. Seasonal Variation in Avian Community Composition in A High-Andean Polylepis (Rosaceae) Forest Fragmentlaspiur22blues7327Оценок пока нет
- Jeb 12188Документ11 страницJeb 12188laspiur22blues7327Оценок пока нет
- Amphisbaena Persephone Sp. NovДокумент12 страницAmphisbaena Persephone Sp. Novlaspiur22blues7327Оценок пока нет
- Sceloporus Virgatus: Comparison of Two Diet Collection Methods in The Lizards, Sceloporus Jarrovii andДокумент4 страницыSceloporus Virgatus: Comparison of Two Diet Collection Methods in The Lizards, Sceloporus Jarrovii andlaspiur22blues7327Оценок пока нет
- J 1095-8649 1996 tb00079 XДокумент13 страницJ 1095-8649 1996 tb00079 Xlaspiur22blues7327Оценок пока нет
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryОт EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryРейтинг: 3.5 из 5 звезд3.5/5 (231)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceОт EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceРейтинг: 4 из 5 звезд4/5 (894)
- The Yellow House: A Memoir (2019 National Book Award Winner)От EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Рейтинг: 4 из 5 звезд4/5 (98)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureОт EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureРейтинг: 4.5 из 5 звезд4.5/5 (474)
- Never Split the Difference: Negotiating As If Your Life Depended On ItОт EverandNever Split the Difference: Negotiating As If Your Life Depended On ItРейтинг: 4.5 из 5 звезд4.5/5 (838)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaОт EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaРейтинг: 4.5 из 5 звезд4.5/5 (265)
- The Emperor of All Maladies: A Biography of CancerОт EverandThe Emperor of All Maladies: A Biography of CancerРейтинг: 4.5 из 5 звезд4.5/5 (271)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersОт EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersРейтинг: 4.5 из 5 звезд4.5/5 (344)
- Team of Rivals: The Political Genius of Abraham LincolnОт EverandTeam of Rivals: The Political Genius of Abraham LincolnРейтинг: 4.5 из 5 звезд4.5/5 (234)
- The Unwinding: An Inner History of the New AmericaОт EverandThe Unwinding: An Inner History of the New AmericaРейтинг: 4 из 5 звезд4/5 (45)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyОт EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyРейтинг: 3.5 из 5 звезд3.5/5 (2219)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreОт EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreРейтинг: 4 из 5 звезд4/5 (1090)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)От EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Рейтинг: 4.5 из 5 звезд4.5/5 (119)
- Man Refined Energetic Seduction - Chris BaleДокумент63 страницыMan Refined Energetic Seduction - Chris BaleSanti Sica0% (2)
- Regular Evaluation Test I Maths+Sc IXДокумент7 страницRegular Evaluation Test I Maths+Sc IXB.L. Indo Anglian Public School, Aurngabad, BiharОценок пока нет
- Life in The Deserts Worksheet 2Документ2 страницыLife in The Deserts Worksheet 2Uma TiwariОценок пока нет
- Population Size, Density, & Dispersal: Demography: Describing Populations and How They ChangeДокумент4 страницыPopulation Size, Density, & Dispersal: Demography: Describing Populations and How They ChangeAllan Jr. Agao-AgaoОценок пока нет
- Organizing A TextДокумент3 страницыOrganizing A TextMercedes Jimenez RomanОценок пока нет
- Development of The Planet EarthДокумент14 страницDevelopment of The Planet EarthHana CpnplnОценок пока нет
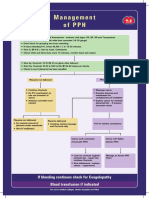
- Management of PPHДокумент1 страницаManagement of PPH098 U.KARTHIK SARAVANA KANTHОценок пока нет
- Comparison of the Fully Automated Urinalysis Analyzers UX-2000 and Cobas 6500Документ10 страницComparison of the Fully Automated Urinalysis Analyzers UX-2000 and Cobas 6500Sethulakshmi PharmacistОценок пока нет
- Characterization of Polydimethylsiloxane (PDMS) PropertiesДокумент13 страницCharacterization of Polydimethylsiloxane (PDMS) PropertiesEsteban ArayaОценок пока нет
- 2 Part 1Документ99 страниц2 Part 1m_luchianОценок пока нет
- The Antibacterial Properties of Isothiocyanates PDFДокумент15 страницThe Antibacterial Properties of Isothiocyanates PDFSasicha DoungsuwanОценок пока нет
- Muscle Energy Techniques ExplainedДокумент2 страницыMuscle Energy Techniques ExplainedAtanu Datta100% (1)
- Egg Quality Manual Web PDFДокумент44 страницыEgg Quality Manual Web PDFVeterinario La EsperanzaОценок пока нет
- 4546-LacOperon InstsДокумент14 страниц4546-LacOperon InstsScribd_is_GreatОценок пока нет
- Dna Finger PrintingДокумент23 страницыDna Finger PrintingSanjay KumarОценок пока нет
- Follicular Dynamics in Bovine and Ovine 1Документ19 страницFollicular Dynamics in Bovine and Ovine 1israr yousafОценок пока нет
- 2nd Periodic Test (Science)Документ1 страница2nd Periodic Test (Science)mosarbas0950% (2)
- Turbidimetry and Nephelometry1Документ3 страницыTurbidimetry and Nephelometry1maxim_crank6101100% (1)
- COT TRF Science G8 - Food ChainДокумент8 страницCOT TRF Science G8 - Food ChainRAMIR BECOYОценок пока нет
- Chapter 2 Medical Terminology Verified AnswersДокумент5 страницChapter 2 Medical Terminology Verified AnswersGregg ProducerОценок пока нет
- UAS Writing For Discourse - 4 Pagi OkДокумент1 страницаUAS Writing For Discourse - 4 Pagi Oksalwa syabinaОценок пока нет
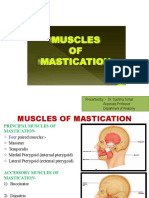
- Presented By:-Dr. Sushma Tomar Associate Professor Department of AnatomyДокумент20 страницPresented By:-Dr. Sushma Tomar Associate Professor Department of AnatomyCristine EchaveОценок пока нет
- DNA TimelineДокумент2 страницыDNA TimelineMinaMilićОценок пока нет
- Microbial Healing of Cracks in ConcreteДокумент15 страницMicrobial Healing of Cracks in ConcreteasdasdasdasОценок пока нет
- Analisis Kualitatif Dan Kuantitatif Kandungan Kimia Dari Ekstrak Heksan, Aseton, Etanol Dan Air Dari Umbi Bawang Putih (Allium Sativum Linn.)Документ11 страницAnalisis Kualitatif Dan Kuantitatif Kandungan Kimia Dari Ekstrak Heksan, Aseton, Etanol Dan Air Dari Umbi Bawang Putih (Allium Sativum Linn.)Tari PratiwiОценок пока нет
- LearnerДокумент7 страницLearnersudhacarhrОценок пока нет
- Biology of Implant OsseointegrationДокумент11 страницBiology of Implant OsseointegrationAmIrОценок пока нет
- Reproductive Biology and Phylogeny of Birds PDFДокумент624 страницыReproductive Biology and Phylogeny of Birds PDFFabricio GZОценок пока нет
- Hemostatic Effects of Peperomia PellucidДокумент41 страницаHemostatic Effects of Peperomia PellucidGina Rose De GuzmanОценок пока нет
- Transpiration POGIL Answer KeyДокумент8 страницTranspiration POGIL Answer KeyJade Tapper50% (2)