Вам также может понравиться
- Codigo Genetico InformeДокумент2 страницыCodigo Genetico InformeLenin Alfaro VidalОценок пока нет
- Microbiologia de Pescados y Mariscos AДокумент41 страницаMicrobiologia de Pescados y Mariscos AJonathan Acuña CordovaОценок пока нет
- Análisis de Competidores y Productos Sustitutos Existentes en El MercadoДокумент7 страницAnálisis de Competidores y Productos Sustitutos Existentes en El MercadoJonathan Acuña Cordova100% (1)
- PreLab 6,7Документ7 страницPreLab 6,7Ana Ceto BritoОценок пока нет
- Alimento para Cuidar y Mantener Un Cabello SaludableДокумент3 страницыAlimento para Cuidar y Mantener Un Cabello SaludableRocio BarretoОценок пока нет
- Musculo Liso (Histologia)Документ3 страницыMusculo Liso (Histologia)Dave KrozОценок пока нет
- Macromoléculas Cuadro ComparativoДокумент4 страницыMacromoléculas Cuadro ComparativoSofia BermudezОценок пока нет
- Tema 3. Tejido Conectivo ChunchunmaruДокумент46 страницTema 3. Tejido Conectivo ChunchunmaruAnto Sevilla15Оценок пока нет
- Factores Que Modifican El Efecto FarmacológicoДокумент11 страницFactores Que Modifican El Efecto FarmacológicoYochito100% (1)
- Beta OxidacionДокумент6 страницBeta OxidacionPao AlmendrasОценок пока нет
- Estructura y Morfología de Los Hongos-ExpoДокумент13 страницEstructura y Morfología de Los Hongos-ExpoNoel Îa Salazar100% (1)
- AVITECNIA - Capà Tulo 16 Procesamiento PDFДокумент19 страницAVITECNIA - Capà Tulo 16 Procesamiento PDFJaqueline Stephanie Ley MartinezОценок пока нет
- Clase 03 - Regeneracion y Reparacion TisularДокумент55 страницClase 03 - Regeneracion y Reparacion TisularVanessa Avila100% (1)
- Anemiaferropenicafull 131010014025 Phpapp02Документ20 страницAnemiaferropenicafull 131010014025 Phpapp02César Obed OlivaresОценок пока нет
- Reparación de MembranasДокумент2 страницыReparación de MembranasAle0% (1)
- Metabolismo de Los MineralesДокумент45 страницMetabolismo de Los MineralesRicardo VallejoОценок пока нет
- Esta Filo CocosДокумент2 страницыEsta Filo CocosdannymalcacamposОценок пока нет
- Cuadro Estratos de Sistema TegumentarioДокумент2 страницыCuadro Estratos de Sistema TegumentarioLourdes GómezОценок пока нет
- 1.introduccion Al MetabolismoДокумент9 страниц1.introduccion Al MetabolismoAndres Felipe Prieto AlarconОценок пока нет
- Cuestionario Determinación de Triglicéridos y ColesterolДокумент4 страницыCuestionario Determinación de Triglicéridos y ColesterolKedwin MontejoОценок пока нет
- Especializaciones de La Región LateralДокумент6 страницEspecializaciones de La Región LateralAmbar Nivar GonzálezОценок пока нет
- 127 Horas AnalisisДокумент2 страницы127 Horas Analisispedro gomezОценок пока нет
- Cangrejo Violáceo-Adam VasquezДокумент4 страницыCangrejo Violáceo-Adam VasquezJoel s100% (1)
- Clase 6 - Estructura de Las ProteínasДокумент46 страницClase 6 - Estructura de Las ProteínasDJ AKRODOK100% (1)
- Polaridad CelularДокумент3 страницыPolaridad CelularHenderson Vasquez ArmasОценок пока нет
- ISOMERASASДокумент9 страницISOMERASASAndia Cch100% (1)
- Kevin Campo 00209473 Informe 2 MacromoleculasДокумент11 страницKevin Campo 00209473 Informe 2 MacromoleculasKevin CampoОценок пока нет
- Disacaridos y Aldosas CetosasДокумент3 страницыDisacaridos y Aldosas Cetosascheshireww0% (1)
- Peroxisomas PDFДокумент3 страницыPeroxisomas PDFJesus Tun100% (1)
- Citología Actual2021Документ111 страницCitología Actual2021Nicol Torrez CabaОценок пока нет
- Capítulo 3 - Aminoácidos y PéptidosДокумент16 страницCapítulo 3 - Aminoácidos y PéptidosmariaОценок пока нет
- Asociación BiológicaДокумент4 страницыAsociación BiológicaMARCO ANTONIO ABARCAОценок пока нет
- Fisiopatología de La PolicitemiaДокумент2 страницыFisiopatología de La PolicitemiaWendy Villacres RiveraОценок пока нет
- Ulcera Peptica Marco Teorico ImprimirДокумент31 страницаUlcera Peptica Marco Teorico ImprimirRouse AguirreОценок пока нет
- Marasmo y KwasshiorkorДокумент1 страницаMarasmo y KwasshiorkorJuliethОценок пока нет
- Practica CiclosisДокумент5 страницPractica CiclosisJhohan Andersson Sanchez YlquimicheОценок пока нет
- Emulsificacion LipidosДокумент16 страницEmulsificacion LipidosJessica CultidОценок пока нет
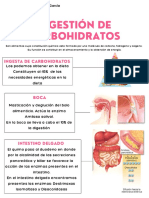
- Digestion de CarbohidratosДокумент3 страницыDigestion de CarbohidratosIvanna HoffmanОценок пока нет
- Proyecto de Investigacion Grupo 6, OsmosisДокумент27 страницProyecto de Investigacion Grupo 6, Osmosisbiounsch_24nilmessi0% (1)
- 2do Ok Biologia - 2okДокумент17 страниц2do Ok Biologia - 2okLuz Elena Armas ChuicaОценок пока нет
- Informe Biologia (Tejidos)Документ24 страницыInforme Biologia (Tejidos)Anderson Vargas0% (2)
- Desnaturalización de ProteínasДокумент4 страницыDesnaturalización de ProteínasJulian PerezОценок пока нет
- Fitoquimica Del Clavo de OlorДокумент5 страницFitoquimica Del Clavo de OlorjoseОценок пока нет
- Cápsula de BealДокумент1 страницаCápsula de BealSergio Barragan100% (3)
- Histología Del Tejido Óseo PDFДокумент24 страницыHistología Del Tejido Óseo PDFKarol Dayana Caro BustamanteОценок пока нет
- Cuadro ComparativoДокумент7 страницCuadro ComparativoperlacastrovОценок пока нет
- Mapa Mental Enzimas Eimar Rivero 201O3Документ1 страницаMapa Mental Enzimas Eimar Rivero 201O3Eimar RiveroОценок пока нет
- Marco Teórico de MicrobiologiaДокумент6 страницMarco Teórico de MicrobiologiaYair Vidal RiveraОценок пока нет
- Ácido Mefenámico 500 MGДокумент2 страницыÁcido Mefenámico 500 MGXimena AndreaОценок пока нет
- Funciones Biológicas de Los 20 AminoácidosДокумент5 страницFunciones Biológicas de Los 20 AminoácidosMadeleine RodriguezОценок пока нет
- SEMANA 8 CitoesqueletoДокумент2 страницыSEMANA 8 CitoesqueletoMartino San MartinОценок пока нет
- Alteraciones Eritrocitarias, Tamaño y ColorДокумент23 страницыAlteraciones Eritrocitarias, Tamaño y ColorHanny Michelle Magaña RaveloОценок пока нет
- Informe de Practica Tejido EpitelialДокумент15 страницInforme de Practica Tejido EpitelialLucia YuquiОценок пока нет
- Cascada de La CoagulaciónДокумент2 страницыCascada de La CoagulaciónMarifer Arbesú100% (1)
- Caso Clinico # 5 Coma HepaticoДокумент10 страницCaso Clinico # 5 Coma HepaticoEr1ck9450% (2)
- Shigella SPPДокумент15 страницShigella SPPANEL CISNEROS FLORESОценок пока нет
- Diapositivas Parásitos en AlimentosДокумент10 страницDiapositivas Parásitos en AlimentosPaola RincónОценок пока нет
- Taenia SaginataДокумент4 страницыTaenia SaginataAngel OrdoñezОценок пока нет
- Mapa Conceptual Cryptococcus NeoformansДокумент2 страницыMapa Conceptual Cryptococcus NeoformansLuis Eduardo Cruz MartínezОценок пока нет
- Cadena de Supervivencia IntrahospitalarioДокумент2 страницыCadena de Supervivencia IntrahospitalarioYENNYFER CAMPOS RIVERAОценок пока нет
- Practicahigadoparablog 120208191849 Phpapp02Документ2 страницыPracticahigadoparablog 120208191849 Phpapp02Piero Angelo Morales AlcaldeОценок пока нет
- IntroducciónДокумент4 страницыIntroducciónAlejandra Marín OlavarríaОценок пока нет
- Desnaturalización de Las ProteínasДокумент7 страницDesnaturalización de Las ProteínasIván Meneses AceroОценок пока нет
- Enzimas PDFДокумент8 страницEnzimas PDFAnonymous KarinaОценок пока нет
- BIOLOGIAДокумент12 страницBIOLOGIAYanixa Vivar MauricioОценок пока нет
- EnzimasДокумент9 страницEnzimasVinicio BermudezОценок пока нет
- Información BiologiaДокумент3 страницыInformación BiologiaEmerson Tiago Camacho DíazОценок пока нет
- RD 20 2015 Digesa SaДокумент2 страницыRD 20 2015 Digesa SaJonathan Acuña CordovaОценок пока нет
- Los ColorantesДокумент29 страницLos ColorantesJonathan Acuña CordovaОценок пока нет
- Aspergillus Oryzae Trabajo FinalДокумент9 страницAspergillus Oryzae Trabajo FinalJonathan Acuña CordovaОценок пока нет
- Estatuto Del Estudiante UniversitarioДокумент7 страницEstatuto Del Estudiante UniversitarioJonathan Acuña CordovaОценок пока нет
- Libro CajaДокумент15 страницLibro CajaJonathan Acuña Cordova0% (1)
- Tesis de Aceite de Pescado 3Документ65 страницTesis de Aceite de Pescado 3Jaime SutizalОценок пока нет
- 1 - Informe de Ensayo - Arroz FortificadoДокумент6 страниц1 - Informe de Ensayo - Arroz FortificadoJorgeRaulValdeiglesiasJaraОценок пока нет
- D4º Act 4-Cyt-U1Документ6 страницD4º Act 4-Cyt-U1Kamila QuirozОценок пока нет
- Catalogo Trasnacional 06 08 19Документ14 страницCatalogo Trasnacional 06 08 19darinelОценок пока нет
- Ensayo de LipidosДокумент13 страницEnsayo de LipidosArturo Hernández Sandoval0% (2)
- El Colágeno Es Una MoléculaДокумент6 страницEl Colágeno Es Una MoléculajosueОценок пока нет
- Sgrasas y AceiteДокумент30 страницSgrasas y AceiteKarime AVОценок пока нет
- LPG Productos Lacteos S.AДокумент44 страницыLPG Productos Lacteos S.APAULA ANDREA LOAIZA ARBOLEDAОценок пока нет
- 4-Lípidos PlasmáticosДокумент22 страницы4-Lípidos PlasmáticosDenia EstradaОценок пока нет
- Principios de Lehninger Del Banco de Pruebas de Bioquímica Capítulo 4Документ9 страницPrincipios de Lehninger Del Banco de Pruebas de Bioquímica Capítulo 4BRYAN ALEXIS ORDOÑEZ RECINOSОценок пока нет
- Harina de TrigoДокумент10 страницHarina de TrigoAna MariaОценок пока нет
- Aminoácidos BetaДокумент2 страницыAminoácidos BetaMarce PerezОценок пока нет
- Carnes AuquenidasДокумент4 страницыCarnes AuquenidasMaykel MorenoОценок пока нет
- 2022 Biología Carbohidratos Clase 1Документ22 страницы2022 Biología Carbohidratos Clase 1SebastianОценок пока нет
- Vitamina EДокумент2 страницыVitamina EzoradonОценок пока нет
- Demuestro Lo Aprendido IbДокумент2 страницыDemuestro Lo Aprendido IbJushepy SandovalОценок пока нет
- Actividad Biomoleculas-Sebastian Coronado Ballesteros - Cultura Fisica, Recreacion y Deporte.Документ2 страницыActividad Biomoleculas-Sebastian Coronado Ballesteros - Cultura Fisica, Recreacion y Deporte.sebastian coronadoОценок пока нет
- PROTEÍNASДокумент45 страницPROTEÍNASPillaca Ugarte Ulises RansesОценок пока нет
- Exámen Final BioquímicaДокумент43 страницыExámen Final BioquímicaEduardo MurilloОценок пока нет
- Examen Biologia Noveno P1Документ2 страницыExamen Biologia Noveno P1Breshier Alfaro-HattumОценок пока нет
- Gua de Productos 2020 USA Digital Vol IIДокумент11 страницGua de Productos 2020 USA Digital Vol IIRoseyronОценок пока нет
- Componentes Qumicos en Las CelulasДокумент20 страницComponentes Qumicos en Las CelulasjoseОценок пока нет
- Aminoacidos CДокумент18 страницAminoacidos CWILFREDO ROMAN PAUCARОценок пока нет
- Biologia Biomoléculas UNAMДокумент15 страницBiologia Biomoléculas UNAMFelipe GarciaОценок пока нет
- Informe de GrasasДокумент5 страницInforme de GrasasDelinda Maria Pereira DurangoОценок пока нет
- Colágenos - Tipos, Composición, Características y Distribución en TejidosДокумент6 страницColágenos - Tipos, Composición, Características y Distribución en TejidosAna SaraviaОценок пока нет