Вам также может понравиться
- The Yellow House: A Memoir (2019 National Book Award Winner)От EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Рейтинг: 4 из 5 звезд4/5 (98)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeОт EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeРейтинг: 4 из 5 звезд4/5 (5794)
- The Little Book of Hygge: Danish Secrets to Happy LivingОт EverandThe Little Book of Hygge: Danish Secrets to Happy LivingРейтинг: 3.5 из 5 звезд3.5/5 (400)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureОт EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureРейтинг: 4.5 из 5 звезд4.5/5 (474)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryОт EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryРейтинг: 3.5 из 5 звезд3.5/5 (231)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceОт EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceРейтинг: 4 из 5 звезд4/5 (895)
- Team of Rivals: The Political Genius of Abraham LincolnОт EverandTeam of Rivals: The Political Genius of Abraham LincolnРейтинг: 4.5 из 5 звезд4.5/5 (234)
- Never Split the Difference: Negotiating As If Your Life Depended On ItОт EverandNever Split the Difference: Negotiating As If Your Life Depended On ItРейтинг: 4.5 из 5 звезд4.5/5 (838)
- The Emperor of All Maladies: A Biography of CancerОт EverandThe Emperor of All Maladies: A Biography of CancerРейтинг: 4.5 из 5 звезд4.5/5 (271)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaОт EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaРейтинг: 4.5 из 5 звезд4.5/5 (266)
- The Unwinding: An Inner History of the New AmericaОт EverandThe Unwinding: An Inner History of the New AmericaРейтинг: 4 из 5 звезд4/5 (45)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersОт EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersРейтинг: 4.5 из 5 звезд4.5/5 (345)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyОт EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyРейтинг: 3.5 из 5 звезд3.5/5 (2259)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreОт EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreРейтинг: 4 из 5 звезд4/5 (1090)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)От EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Рейтинг: 4.5 из 5 звезд4.5/5 (121)
- ECO-321 Development Economics: Instructor Name: Syeda Nida RazaДокумент10 страницECO-321 Development Economics: Instructor Name: Syeda Nida RazaLaiba MalikОценок пока нет
- Photosynthesis Knowledge OrganiserДокумент1 страницаPhotosynthesis Knowledge Organiserapi-422428700Оценок пока нет
- Network Access Control Quiz3 PDFДокумент2 страницыNetwork Access Control Quiz3 PDFDaljeet SinghОценок пока нет
- R. Raghunandanan - 6Документ48 страницR. Raghunandanan - 6fitrohtin hidayatiОценок пока нет
- EarthmattersДокумент7 страницEarthmattersfeafvaevsОценок пока нет
- English Quarterly TestДокумент3 страницыEnglish Quarterly TestEdmon FabregasОценок пока нет
- H.influenzae Modified 2012Документ12 страницH.influenzae Modified 2012MoonAIRОценок пока нет
- Study On Optimization and Performance of Biological Enhanced Activated Sludge Process For Pharmaceutical Wastewater TreatmentДокумент9 страницStudy On Optimization and Performance of Biological Enhanced Activated Sludge Process For Pharmaceutical Wastewater TreatmentVijaya GosuОценок пока нет
- Air Pollution Emissions 2008-2018 From Australian Coal Mining: Implications For Public and Occupational HealthДокумент11 страницAir Pollution Emissions 2008-2018 From Australian Coal Mining: Implications For Public and Occupational HealthMaria Stephany CalisayaОценок пока нет
- Stats Review CH 1-6Документ15 страницStats Review CH 1-6Megha BanerjeeОценок пока нет
- Nursing Care Plan ConstipationДокумент3 страницыNursing Care Plan Constipationbmrose3783% (12)
- MODEL 1332 MODEL 1432: Slide LengthДокумент8 страницMODEL 1332 MODEL 1432: Slide LengthFabian FragosoОценок пока нет
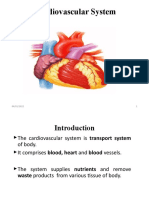
- Cardiovascular SystemДокумент47 страницCardiovascular SystemAtnasiya GadisaОценок пока нет
- Akin To PityДокумент356 страницAkin To PityPaul StewartОценок пока нет
- GASESДокумент55 страницGASESja_QuinineОценок пока нет
- 2 Avaliação Edros 2023 - 7º Ano - ProvaДокумент32 страницы2 Avaliação Edros 2023 - 7º Ano - Provaleandro costaОценок пока нет
- Question Bank Chemistry (B.Tech.) : Solid StateДокумент10 страницQuestion Bank Chemistry (B.Tech.) : Solid StatenraiinОценок пока нет
- Industrial Attachment ReportДокумент34 страницыIndustrial Attachment ReportOtsile Charisma Otsile Saq100% (1)
- 30356-Article Text-56848-1-10-20210201Документ14 страниц30356-Article Text-56848-1-10-20210201Mel FaithОценок пока нет
- Chapter 23Документ9 страницChapter 23Trixie Myr AndoyОценок пока нет
- Writing The Motherland From The DiasporaДокумент23 страницыWriting The Motherland From The DiasporaIfeoluwa Watson100% (1)
- Dyestone Blue MX SDS SA-0186-01Документ5 страницDyestone Blue MX SDS SA-0186-01gede aris prayoga mahardikaОценок пока нет
- Sim Medium 2 (English)Документ2 страницыSim Medium 2 (English)TheLobitoОценок пока нет
- UntitledДокумент2 страницыUntitledapi-236961637Оценок пока нет
- Water Quantity Estimation PDFДокумент3 страницыWater Quantity Estimation PDFOladunni AfolabiОценок пока нет
- 5219-PRP Support Tool CounsellingДокумент10 страниц5219-PRP Support Tool CounsellingRodhi AnshariОценок пока нет
- PACKET 1 - Exercises and ActivitiesДокумент7 страницPACKET 1 - Exercises and ActivitiesPrantsir Pernes100% (2)
- SCL NotesДокумент4 страницыSCL NotesmayaОценок пока нет
- Public Health Interventions: Applications For Public Health Nursing PracticeДокумент249 страницPublic Health Interventions: Applications For Public Health Nursing PracticeJemimah AdaclogОценок пока нет
- Review Factors Contributing To Medication Errors: A Literature ReviewДокумент9 страницReview Factors Contributing To Medication Errors: A Literature Reviewsoul_0602Оценок пока нет