Вам также может понравиться
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryОт EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryРейтинг: 3.5 из 5 звезд3.5/5 (231)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)От EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Рейтинг: 4.5 из 5 звезд4.5/5 (121)
- Never Split the Difference: Negotiating As If Your Life Depended On ItОт EverandNever Split the Difference: Negotiating As If Your Life Depended On ItРейтинг: 4.5 из 5 звезд4.5/5 (838)
- The Little Book of Hygge: Danish Secrets to Happy LivingОт EverandThe Little Book of Hygge: Danish Secrets to Happy LivingРейтинг: 3.5 из 5 звезд3.5/5 (400)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaОт EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaРейтинг: 4.5 из 5 звезд4.5/5 (266)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeОт EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeРейтинг: 4 из 5 звезд4/5 (5795)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreОт EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreРейтинг: 4 из 5 звезд4/5 (1090)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyОт EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyРейтинг: 3.5 из 5 звезд3.5/5 (2259)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersОт EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersРейтинг: 4.5 из 5 звезд4.5/5 (345)
- The Emperor of All Maladies: A Biography of CancerОт EverandThe Emperor of All Maladies: A Biography of CancerРейтинг: 4.5 из 5 звезд4.5/5 (271)
- Team of Rivals: The Political Genius of Abraham LincolnОт EverandTeam of Rivals: The Political Genius of Abraham LincolnРейтинг: 4.5 из 5 звезд4.5/5 (234)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceОт EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceРейтинг: 4 из 5 звезд4/5 (895)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureОт EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureРейтинг: 4.5 из 5 звезд4.5/5 (474)
- The Yellow House: A Memoir (2019 National Book Award Winner)От EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Рейтинг: 4 из 5 звезд4/5 (98)
- The Unwinding: An Inner History of the New AmericaОт EverandThe Unwinding: An Inner History of the New AmericaРейтинг: 4 из 5 звезд4/5 (45)
- Photosynthesis Worksheet AnДокумент3 страницыPhotosynthesis Worksheet AnMa Leslynne100% (1)
- How To Determine The Age and Sex of A VictimДокумент27 страницHow To Determine The Age and Sex of A VictimpoovenmoonoosamyОценок пока нет
- AppliedДокумент619 страницAppliedLau GVОценок пока нет
- Quarter 1 - Module 7 S9LT-Ig-j-31: PhotosynthesisДокумент9 страницQuarter 1 - Module 7 S9LT-Ig-j-31: PhotosynthesisJulia Clarisse Alanza BallonОценок пока нет
- CALVIN CycleДокумент13 страницCALVIN CycleDaphne CuaresmaОценок пока нет
- CapsuleДокумент46 страницCapsulepoovenmoonoosamyОценок пока нет
- Group Number: 02 Course: Physical Pharmacy Course Code: 311Документ70 страницGroup Number: 02 Course: Physical Pharmacy Course Code: 311poovenmoonoosamyОценок пока нет
- Distinguishing Between Human and Non-Human BonesДокумент70 страницDistinguishing Between Human and Non-Human BonespoovenmoonoosamyОценок пока нет
- Vertebrate Development: Biology II: Form and FunctionДокумент41 страницаVertebrate Development: Biology II: Form and FunctionpoovenmoonoosamyОценок пока нет
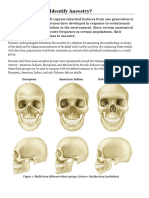
- Activity: Can You Identify Ancestry?: European American Indian Sub-Saharan AfricanДокумент5 страницActivity: Can You Identify Ancestry?: European American Indian Sub-Saharan AfricanpoovenmoonoosamyОценок пока нет
- Sun ScreenДокумент8 страницSun ScreenJai MurugeshОценок пока нет
- TMP 8 FD1Документ38 страницTMP 8 FD1FrontiersОценок пока нет
- FGHДокумент231 страницаFGHWalid Ebid Elgammal100% (1)
- A Plant Fisio IntroДокумент2 страницыA Plant Fisio IntroamirahrazaliОценок пока нет
- GenBio1 Week 1Документ14 страницGenBio1 Week 1ALIÑO, AHZILLE P.Оценок пока нет
- Photosynthesis Coloring 1Документ2 страницыPhotosynthesis Coloring 1api-2608174140% (1)
- Photosyn-Thesis: Calvin Cycle: Reporters: Group 4Документ11 страницPhotosyn-Thesis: Calvin Cycle: Reporters: Group 4John Allen GuisdanОценок пока нет
- CH 12Документ26 страницCH 12Yogesh MudgalОценок пока нет
- Engl, Reiser - 2020Документ8 страницEngl, Reiser - 2020franciscaОценок пока нет
- Chapter 10 BioДокумент2 страницыChapter 10 BioPaul Meyer75% (8)
- BIOC 2061 Tutorial 8Документ7 страницBIOC 2061 Tutorial 8Tishelle McCarthyОценок пока нет
- Photosynthesis in Higher Plants - Shobhit NirwanДокумент21 страницаPhotosynthesis in Higher Plants - Shobhit NirwanVraj M Barot100% (1)
- A2 Photosynthesis Homework StudyДокумент3 страницыA2 Photosynthesis Homework StudystjustinsОценок пока нет
- Photosynthesis Summary By: Abdul Majid HasaniДокумент12 страницPhotosynthesis Summary By: Abdul Majid HasaniSani Stickman AxcОценок пока нет
- 5A-IAL - ExamwizardДокумент27 страниц5A-IAL - ExamwizarddualipaОценок пока нет
- I. Photosynthesis (Light and Dark Reaction) : Quarter 2 / Semester 1, Week 2Документ6 страницI. Photosynthesis (Light and Dark Reaction) : Quarter 2 / Semester 1, Week 2Monica SolomonОценок пока нет
- Photolysis of WaterДокумент4 страницыPhotolysis of WaterDharmendra SinghОценок пока нет
- Photosynthesis PDFДокумент22 страницыPhotosynthesis PDFbhaskar rayОценок пока нет
- BSC PDFДокумент69 страницBSC PDFNeha SharmaОценок пока нет
- PHOTOSYNTHESISДокумент32 страницыPHOTOSYNTHESISArlene SantosОценок пока нет
- Photosynthesis in Higher PlantsДокумент8 страницPhotosynthesis in Higher PlantsAditya VenkatОценок пока нет
- Photosynthesis - Simple English Wikipedia The Free EncyclopediaДокумент3 страницыPhotosynthesis - Simple English Wikipedia The Free Encyclopediaapi-253604375Оценок пока нет
- 13 - Photosynthesis AQA BookletДокумент28 страниц13 - Photosynthesis AQA BookletSevilay CaferogluОценок пока нет
- Apaharan (1,2) - Qazi Anwar HossainДокумент386 страницApaharan (1,2) - Qazi Anwar Hossainhsr1708Оценок пока нет
- Photosynthesis Hhmi BiointeractiveДокумент7 страницPhotosynthesis Hhmi BiointeractivemariaОценок пока нет
- 2022-Synoxyl® AZ BrochureДокумент12 страниц2022-Synoxyl® AZ BrochureMihaela Simona IsacОценок пока нет