Вам также может понравиться
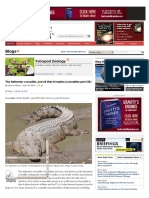
- Casquehead Iguana PDFДокумент3 страницыCasquehead Iguana PDFJohn GamesbyОценок пока нет
- Keys To The Nematode Parasites of Vertebrates, Archival Volume (VetBooks - Ir)Документ475 страницKeys To The Nematode Parasites of Vertebrates, Archival Volume (VetBooks - Ir)YoshioОценок пока нет
- Cracraft & Donoghue (Eds.) - Assembling The Tree of Life (2004) PDFДокумент594 страницыCracraft & Donoghue (Eds.) - Assembling The Tree of Life (2004) PDFAtman0% (1)
- Acta Tropica: P. Coelho, P. Sousa, D.J. Harris, A. Van Der MeijdenДокумент9 страницActa Tropica: P. Coelho, P. Sousa, D.J. Harris, A. Van Der MeijdenInes AquariusОценок пока нет
- Phylogeography of Androctonus Species (Scorpiones: Buthidae) in Tunisia: Diagnostic Characters For Linking Species To ScorpionismДокумент9 страницPhylogeography of Androctonus Species (Scorpiones: Buthidae) in Tunisia: Diagnostic Characters For Linking Species To ScorpionismInes AquariusОценок пока нет
- New and Additional Records For The Ant Fauna (Hymenoptera, Formicidae) of MoroccoДокумент10 страницNew and Additional Records For The Ant Fauna (Hymenoptera, Formicidae) of Moroccodavid fernandezОценок пока нет
- Clave Gráfica de Adultos de Los Geeros Hematofagos de Ceratopogonidae Presentes en La Region NeotropicalДокумент15 страницClave Gráfica de Adultos de Los Geeros Hematofagos de Ceratopogonidae Presentes en La Region NeotropicalLuis Paulo Costa CarvalhoОценок пока нет
- Orugas EcuadorДокумент15 страницOrugas EcuadorJulian AlzateОценок пока нет
- Genetic Diversity of Ribosomal RNA Internal Transcribed Spacer Sequences in Cutaneous LeishmaniasisДокумент6 страницGenetic Diversity of Ribosomal RNA Internal Transcribed Spacer Sequences in Cutaneous LeishmaniasisFlor EsthefanyОценок пока нет
- Tadpole, Oophagy, Advertisement Call, and Geographic Distribution of Aparasphenodon Arapapa Pimenta, Napoli and Haddad 2009 (Anura, Hylidae)Документ6 страницTadpole, Oophagy, Advertisement Call, and Geographic Distribution of Aparasphenodon Arapapa Pimenta, Napoli and Haddad 2009 (Anura, Hylidae)Guilherme SousaОценок пока нет
- Karyotypes of The Mammals of Turkey and Neighbouring Regions: A ReviewДокумент63 страницыKaryotypes of The Mammals of Turkey and Neighbouring Regions: A ReviewtigranhrptnОценок пока нет
- Hojun Song 2018Документ25 страницHojun Song 2018johanna abrilОценок пока нет
- Wasko 2010Документ12 страницWasko 2010NacorОценок пока нет
- 174783-Article Text-447346-1-10-20180719Документ4 страницы174783-Article Text-447346-1-10-20180719Richard GladОценок пока нет
- First Report of Xiphinema Ifacolum Luc, 1961 (Dorylaimida: Longidoridae) From NigeriaДокумент5 страницFirst Report of Xiphinema Ifacolum Luc, 1961 (Dorylaimida: Longidoridae) From NigeriaTesleem BelloОценок пока нет
- 2010 Occurrence and Infection Dynamics of Anisakid Larvae in Scomber ScombrusДокумент6 страниц2010 Occurrence and Infection Dynamics of Anisakid Larvae in Scomber ScombrusIlham RoyyanОценок пока нет
- Widespread Occurrence of The Amphibian CДокумент8 страницWidespread Occurrence of The Amphibian Cabeje kassieОценок пока нет
- Trogidae) in Exotic and Native Environments of BrazilДокумент4 страницыTrogidae) in Exotic and Native Environments of Brazilflechtma1466Оценок пока нет
- Sanchez-Piñero Et Al. Use of Quercus by Thorectes Species. Afric - EntomolДокумент8 страницSanchez-Piñero Et Al. Use of Quercus by Thorectes Species. Afric - EntomolMarina Trillo GabaldonОценок пока нет
- J. Med. Entomol. 1-8 (2015) DOI: 10.1093/jme/tjv127: Ampling Istribution IspersalДокумент8 страницJ. Med. Entomol. 1-8 (2015) DOI: 10.1093/jme/tjv127: Ampling Istribution IspersalAna MaríaОценок пока нет
- 1 s2.0 S0022201122001598 MainДокумент11 страниц1 s2.0 S0022201122001598 MainshikingstarОценок пока нет
- Phylogeny, Biogeography, and Diversification of Barn Owls (Aves: Strigiformes)Документ15 страницPhylogeny, Biogeography, and Diversification of Barn Owls (Aves: Strigiformes)Oner ManОценок пока нет
- RouagETALAfrJHerp572103 113Документ12 страницRouagETALAfrJHerp572103 113Mohamed MebarkiОценок пока нет
- A Pictorial Key For Identification of The Hoverflies (Diptera: Syrphidae) of The Madeira ArchipelagoДокумент62 страницыA Pictorial Key For Identification of The Hoverflies (Diptera: Syrphidae) of The Madeira ArchipelagoJorge Alonso HerreroОценок пока нет
- Diversity of Scorpions (Chelicerata: Arachnida) in The Atlantic Forest in Pernambuco, Northeastern BrazilДокумент5 страницDiversity of Scorpions (Chelicerata: Arachnida) in The Atlantic Forest in Pernambuco, Northeastern BrazilGonzalo Ramírez GalánОценок пока нет
- Aspects of The Emergence Ecology of The Regionally Endangered Coenagrion Mercuriale Odonata Coenagrionidae in Northeast AlgeriaДокумент8 страницAspects of The Emergence Ecology of The Regionally Endangered Coenagrion Mercuriale Odonata Coenagrionidae in Northeast AlgeriaMohamed Khalil MellalОценок пока нет
- TMP 5 AA9Документ8 страницTMP 5 AA9FrontiersОценок пока нет
- Animals: Morphometric Study of Mus Musculus, Rattus Norvegicus, and Rattus Rattus in QatarДокумент15 страницAnimals: Morphometric Study of Mus Musculus, Rattus Norvegicus, and Rattus Rattus in Qatarjlee_296737Оценок пока нет
- Fouquet 2007Документ16 страницFouquet 2007Carol AraújoОценок пока нет
- De Meyeretal - 2012 - TephritidaecomoresДокумент10 страницDe Meyeretal - 2012 - Tephritidaecomoresperivou agoulaОценок пока нет
- Httner 2008Документ8 страницHttner 2008gabriel avilaОценок пока нет
- An Overview of The Dragonflies and Damselflies of The Maltese Islands (Central Mediterranean) (Odonata)Документ123 страницыAn Overview of The Dragonflies and Damselflies of The Maltese Islands (Central Mediterranean) (Odonata)Ram LeeОценок пока нет
- Aegilops Vavilovii: Taxonomy, Morphology and Palynology of (Zhuk.) Chennav. (Poaceae: Triticeae)Документ9 страницAegilops Vavilovii: Taxonomy, Morphology and Palynology of (Zhuk.) Chennav. (Poaceae: Triticeae)Evren CabiОценок пока нет
- 115 - Euthycaelus Cunampia, Melloina Pacifica, Neischnocolus MecanaДокумент42 страницы115 - Euthycaelus Cunampia, Melloina Pacifica, Neischnocolus MecanaVulpesОценок пока нет
- Linares Et Al. 2016 - (First Report On Predation of Adult Anurans by Odonata Larvae)Документ3 страницыLinares Et Al. 2016 - (First Report On Predation of Adult Anurans by Odonata Larvae)Antônio LinaresОценок пока нет
- Description and Nesting Biology of Three New Species of Neotropical Silk Wasp Hymenoptera Apoidea Pemphredoninae MicrostigmusДокумент19 страницDescription and Nesting Biology of Three New Species of Neotropical Silk Wasp Hymenoptera Apoidea Pemphredoninae MicrostigmusMarcus PrestesОценок пока нет
- Ramos Etal 2022 Evolution Andrenine BeesДокумент11 страницRamos Etal 2022 Evolution Andrenine BeesTayaneFebroneОценок пока нет
- 44023-Article Text-138036-150338-10-20181227Документ11 страниц44023-Article Text-138036-150338-10-20181227Lara Caroline Carvalho RezendeОценок пока нет
- Murakami 2015Документ8 страницMurakami 2015FenixPPОценок пока нет
- Anopheles: Morphological Identification of Malaria Vectors Within Species in Parts of Kano State, NigeriaДокумент4 страницыAnopheles: Morphological Identification of Malaria Vectors Within Species in Parts of Kano State, NigeriaDendy AriezaputraОценок пока нет
- Weinbeer - Kalko. 2004. Morphological Characterictics Predict Alternate Foraging Strategy and Microhabitat Selection in The Orange-Bellied BatДокумент8 страницWeinbeer - Kalko. 2004. Morphological Characterictics Predict Alternate Foraging Strategy and Microhabitat Selection in The Orange-Bellied BatJ E Fernando CernaОценок пока нет
- 230953-Article Text-560517-1-10-20220831Документ14 страниц230953-Article Text-560517-1-10-20220831fireworkОценок пока нет
- Supérvivencia y LogebidadДокумент10 страницSupérvivencia y LogebidadDaniel VargasОценок пока нет
- Gull (Larus Michahellis) in The Aguéli Island of Réghaia, AlgeriaДокумент8 страницGull (Larus Michahellis) in The Aguéli Island of Réghaia, AlgeriaTJPRC PublicationsОценок пока нет
- 1 s2.0 S235198942200213X MainДокумент12 страниц1 s2.0 S235198942200213X MainSUCUITANA CCAIHUARI ANGELI ESTHERОценок пока нет
- Systematic Revision of The Living African Slender-Snouted CrocodilesДокумент43 страницыSystematic Revision of The Living African Slender-Snouted CrocodilesFelipe MesVasОценок пока нет
- 9e58 PDFДокумент10 страниц9e58 PDFcarlos javierОценок пока нет
- De Meyeretal 2013 WestafricanfruitfliesДокумент17 страницDe Meyeretal 2013 Westafricanfruitfliesperivou agoulaОценок пока нет
- TMP 7 B88Документ7 страницTMP 7 B88FrontiersОценок пока нет
- Apis Florea MorphometricsДокумент19 страницApis Florea MorphometricsprabhudevmvОценок пока нет
- 1 s2.0 S0048969723018752 MainДокумент13 страниц1 s2.0 S0048969723018752 MainPedroHenriqueОценок пока нет
- Hard Ticks Parasitizing European Ground Squirrel Spermophilus Citellus (L., 1766) (Rodentia Sciuridae) in SerbiaДокумент8 страницHard Ticks Parasitizing European Ground Squirrel Spermophilus Citellus (L., 1766) (Rodentia Sciuridae) in SerbiaRatko SukaraОценок пока нет
- Hubert Et Al 2019Документ13 страницHubert Et Al 2019Livia TanjungОценок пока нет
- Good Man 2015Документ9 страницGood Man 2015PochoОценок пока нет
- Ectoparasitic Flies (Diptera: Streblidae) On Bats (Mammalia: Chiroptera) From A Private Natural Heritage Reserve in Southeastern BrazilДокумент6 страницEctoparasitic Flies (Diptera: Streblidae) On Bats (Mammalia: Chiroptera) From A Private Natural Heritage Reserve in Southeastern BrazilDaniel SantosОценок пока нет
- Morphometric and Genetic Variations of Three Grouper Species (GenusДокумент7 страницMorphometric and Genetic Variations of Three Grouper Species (Genusmochammad zamroniОценок пока нет
- A Survey of Tropical Ea - Rthworms: Taxonomy, Biogeography and Environmental PlasticityДокумент28 страницA Survey of Tropical Ea - Rthworms: Taxonomy, Biogeography and Environmental Plasticityvipulsingh161Оценок пока нет
- Artículo de RenacuajosДокумент9 страницArtículo de RenacuajosAndres Felipe BermudezОценок пока нет
- A New, Morphologically Cryptic Species of Fanged Frog, Genus Limnonectes From Mindoro Island, Central PhilippinesДокумент24 страницыA New, Morphologically Cryptic Species of Fanged Frog, Genus Limnonectes From Mindoro Island, Central Philippinesdave412Оценок пока нет
- Motta Tavaresetal.2016 EcologyofPhyllodytesluteolusДокумент12 страницMotta Tavaresetal.2016 EcologyofPhyllodytesluteolusvanessa e bruno becaciciОценок пока нет
- Factors Influencing The Level of Infestation of Ixodes Ricinus (Acari-Ixodidae) On Lacerta Agilis and Zootoca Vivipara (Squamata-Lacertidae)Документ9 страницFactors Influencing The Level of Infestation of Ixodes Ricinus (Acari-Ixodidae) On Lacerta Agilis and Zootoca Vivipara (Squamata-Lacertidae)KlausОценок пока нет
- Molecular Ecology - 2020 - Soghigian - Genetic Evidence For The Origin of Aedes Aegypti The Yellow Fever Mosquito in TheДокумент14 страницMolecular Ecology - 2020 - Soghigian - Genetic Evidence For The Origin of Aedes Aegypti The Yellow Fever Mosquito in TheGabriella StefanieОценок пока нет
- Saltwater Crocodile-Crocodiles Part IIIДокумент21 страницаSaltwater Crocodile-Crocodiles Part IIIJohn GamesbyОценок пока нет
- HingebackTortoise PDFДокумент4 страницыHingebackTortoise PDFJohn GamesbyОценок пока нет
- Giant Snakes of Bygone Days PDFДокумент11 страницGiant Snakes of Bygone Days PDFJohn GamesbyОценок пока нет
- Dwarf Boas of The Caribbean PDFДокумент5 страницDwarf Boas of The Caribbean PDFJohn GamesbyОценок пока нет
- Morphological Correlates of Incipient Ar PDFДокумент10 страницMorphological Correlates of Incipient Ar PDFJohn GamesbyОценок пока нет
- Authors Metrics Comments Related ContentДокумент15 страницAuthors Metrics Comments Related ContentJohn GamesbyОценок пока нет
- Crocodile Empire Crocodiles Part 1 PDFДокумент21 страницаCrocodile Empire Crocodiles Part 1 PDFJohn GamesbyОценок пока нет
- An Introduction To Wolf SpidersДокумент4 страницыAn Introduction To Wolf SpidersJohn GamesbyОценок пока нет
- Calabar Python PDFДокумент3 страницыCalabar Python PDFJohn GamesbyОценок пока нет
- A Paper On Cactaceae, Lobivia Ferox A New Record in Chilean FloraДокумент8 страницA Paper On Cactaceae, Lobivia Ferox A New Record in Chilean FloraJohn GamesbyОценок пока нет
- Care Sheet - Pterinochilus MurinusДокумент2 страницыCare Sheet - Pterinochilus MurinusJohn GamesbyОценок пока нет
- Flora of Peak Jabre, Paraíba, Brazilcactaceae - JussДокумент6 страницFlora of Peak Jabre, Paraíba, Brazilcactaceae - JussJohn GamesbyОценок пока нет
- Care Sheet - Plated LizardsДокумент3 страницыCare Sheet - Plated LizardsJohn GamesbyОценок пока нет
- Carrasco Etal 2019 ZootaxaДокумент45 страницCarrasco Etal 2019 Zootaxajoao victorОценок пока нет
- Mscbot 504Документ277 страницMscbot 504Ankit KumarОценок пока нет
- Bioinformatics Assignment Topic: Phylogenetics Analysis SoftwaresДокумент12 страницBioinformatics Assignment Topic: Phylogenetics Analysis Softwaresnidhi teotiaОценок пока нет
- Incongruence Between Cladistic and Taxonomic Systems PDFДокумент8 страницIncongruence Between Cladistic and Taxonomic Systems PDFJavi CastilloОценок пока нет
- KERAGAMAN GENETIK GURITA Octopus Cyanea (LINNAEUS 1758) PAPUAДокумент9 страницKERAGAMAN GENETIK GURITA Octopus Cyanea (LINNAEUS 1758) PAPUAJen SajaОценок пока нет
- Original PDF Molecular and Genome Evolution PDFДокумент41 страницаOriginal PDF Molecular and Genome Evolution PDFtiffany.gonzalez180100% (39)
- Chapter 13 AtelesДокумент43 страницыChapter 13 AtelesManuel RuizОценок пока нет
- TaxonДокумент200 страницTaxonSimion MehedintiОценок пока нет
- Guo 2020Документ10 страницGuo 2020Mohamad Saeful HidayatОценок пока нет
- Paz-Sedano GoniodorididaeДокумент11 страницPaz-Sedano GoniodorididaetemarioguardiaОценок пока нет
- PCR ProtocolsДокумент4 страницыPCR ProtocolsSalvador Emmanuel Sanchez CuapioОценок пока нет
- Grade 12: Systematics Based On Evolutionary Relationships: Cladistics and PhylogenyДокумент14 страницGrade 12: Systematics Based On Evolutionary Relationships: Cladistics and PhylogenyDrew MalubagОценок пока нет
- It Semvii and Viii RC 2016-17-1Документ58 страницIt Semvii and Viii RC 2016-17-1urvesh vernekarОценок пока нет
- Crouch 2009Документ9 страницCrouch 2009ClaudiaFariaОценок пока нет
- Deepthi Scodab PosterДокумент1 страницаDeepthi Scodab PosterMuralidhar MettaОценок пока нет
- Introduction To SystematicsДокумент56 страницIntroduction To SystematicsVeronica MedranoОценок пока нет
- SIR-V6 (4) 3 - FormattedДокумент20 страницSIR-V6 (4) 3 - Formattedaqsa sattarОценок пока нет
- History of Animal Taxonomy v2Документ5 страницHistory of Animal Taxonomy v2Cristie SumbillaОценок пока нет
- Phylogenetic AnalysisДокумент6 страницPhylogenetic AnalysisSheila Mae AramanОценок пока нет
- 1 PBДокумент7 страниц1 PButama 3002Оценок пока нет
- Fungal Identification Using Molecular Tools A Primer For The NaturalДокумент15 страницFungal Identification Using Molecular Tools A Primer For The NaturalSander BroersОценок пока нет
- Cbe 513Документ11 страницCbe 513duverney.gaviriaОценок пока нет
- Phytopathogenic Dothideomycetes (PDFDrive)Документ421 страницаPhytopathogenic Dothideomycetes (PDFDrive)Happy CatsОценок пока нет
- (Methods in Molecular Biology 856) Christian N. K. Anderson, Liang Liu, Dennis Pearl, Scott V. Edwards (auth.), Maria Anisimova (eds.)-Evolutionary Genomics_ Statistical and Computational Method (1).pdfДокумент552 страницы(Methods in Molecular Biology 856) Christian N. K. Anderson, Liang Liu, Dennis Pearl, Scott V. Edwards (auth.), Maria Anisimova (eds.)-Evolutionary Genomics_ Statistical and Computational Method (1).pdfHuGo Castelán SОценок пока нет
- Wen, J. Et Al. (2013) - Biogeography, Where Do We Go From HereДокумент16 страницWen, J. Et Al. (2013) - Biogeography, Where Do We Go From HereJuan David Rodriguez HurtadoОценок пока нет
- OGDEN Et Al-2009-Systematic EntomologyДокумент19 страницOGDEN Et Al-2009-Systematic EntomologyJulianFelipePeñaОценок пока нет
- Study Unit 5: Evolutionary Genetics: 26.1 Organisms Evolve Through Genetic Change Taking Place Within PopulationsДокумент10 страницStudy Unit 5: Evolutionary Genetics: 26.1 Organisms Evolve Through Genetic Change Taking Place Within PopulationsKayleigh MastersОценок пока нет
- A Phylogenetic Assessment of Pholiota and The New Genus PyrrhulomycesДокумент23 страницыA Phylogenetic Assessment of Pholiota and The New Genus PyrrhulomycesFernando FabriniОценок пока нет