Вам также может понравиться
- The Yellow House: A Memoir (2019 National Book Award Winner)От EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Рейтинг: 4 из 5 звезд4/5 (98)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeОт EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeРейтинг: 4 из 5 звезд4/5 (5795)
- Aston-Jones & Cohen (ARN 05)Документ50 страницAston-Jones & Cohen (ARN 05)Rehaan FayazОценок пока нет
- 01 Basics of Computers PDFДокумент19 страниц01 Basics of Computers PDFRehaan FayazОценок пока нет
- FAQs ImportДокумент10 страницFAQs ImportRehaan FayazОценок пока нет
- Sustainable Development Goals (SDGS)Документ4 страницыSustainable Development Goals (SDGS)Rehaan FayazОценок пока нет
- Government Schemes in India: Scheme Name Launch Sector DetailsДокумент8 страницGovernment Schemes in India: Scheme Name Launch Sector DetailsRehaan FayazОценок пока нет
- 1st Sssion On Regulations23052018byFSSAIДокумент20 страниц1st Sssion On Regulations23052018byFSSAIRehaan FayazОценок пока нет
- International Headquarters Organisation Headquarters Head/Secretary GeneralДокумент7 страницInternational Headquarters Organisation Headquarters Head/Secretary GeneralRehaan FayazОценок пока нет
- Women and Armed Conflict: Widows in Kashmir': International Journal of Sociology and AnthropologyДокумент8 страницWomen and Armed Conflict: Widows in Kashmir': International Journal of Sociology and AnthropologyRehaan FayazОценок пока нет
- Review Article: Extracellular Vesicles and Their Convergence With Viral PathwaysДокумент13 страницReview Article: Extracellular Vesicles and Their Convergence With Viral PathwaysRehaan FayazОценок пока нет
- DBTBaramulla PDFДокумент24 страницыDBTBaramulla PDFRehaan FayazОценок пока нет
- UPSC Prelims Result 16Документ43 страницыUPSC Prelims Result 16rafique41cbОценок пока нет
- Issues and Opinions in Nutrition: Utility and Importance of Gene Knockout Animals For Nutritional and Metabolic ResearchДокумент6 страницIssues and Opinions in Nutrition: Utility and Importance of Gene Knockout Animals For Nutritional and Metabolic ResearchRehaan FayazОценок пока нет
- VLPO, TMN, LC InteractionДокумент1 страницаVLPO, TMN, LC InteractionRehaan FayazОценок пока нет
- J. Biol. Chem.-2004-Askwith-18296-305Документ11 страницJ. Biol. Chem.-2004-Askwith-18296-305Rehaan FayazОценок пока нет
- NN 2344Документ8 страницNN 2344Rehaan FayazОценок пока нет
- Test FormДокумент2 страницыTest FormRehaan FayazОценок пока нет
- PHD 2Документ11 страницPHD 2Rehaan FayazОценок пока нет
- Indian GeographyДокумент378 страницIndian GeographyRehaan Fayaz100% (1)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureОт EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureРейтинг: 4.5 из 5 звезд4.5/5 (474)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryОт EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryРейтинг: 3.5 из 5 звезд3.5/5 (231)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceОт EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceРейтинг: 4 из 5 звезд4/5 (895)
- Never Split the Difference: Negotiating As If Your Life Depended On ItОт EverandNever Split the Difference: Negotiating As If Your Life Depended On ItРейтинг: 4.5 из 5 звезд4.5/5 (838)
- The Little Book of Hygge: Danish Secrets to Happy LivingОт EverandThe Little Book of Hygge: Danish Secrets to Happy LivingРейтинг: 3.5 из 5 звезд3.5/5 (400)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersОт EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersРейтинг: 4.5 из 5 звезд4.5/5 (345)
- The Unwinding: An Inner History of the New AmericaОт EverandThe Unwinding: An Inner History of the New AmericaРейтинг: 4 из 5 звезд4/5 (45)
- Team of Rivals: The Political Genius of Abraham LincolnОт EverandTeam of Rivals: The Political Genius of Abraham LincolnРейтинг: 4.5 из 5 звезд4.5/5 (234)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyОт EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyРейтинг: 3.5 из 5 звезд3.5/5 (2259)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaОт EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaРейтинг: 4.5 из 5 звезд4.5/5 (266)
- The Emperor of All Maladies: A Biography of CancerОт EverandThe Emperor of All Maladies: A Biography of CancerРейтинг: 4.5 из 5 звезд4.5/5 (271)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreОт EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreРейтинг: 4 из 5 звезд4/5 (1090)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)От EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Рейтинг: 4.5 из 5 звезд4.5/5 (121)
- Semi-Conductor Laboratory - MohaliДокумент4 страницыSemi-Conductor Laboratory - MohalimanjnderОценок пока нет
- Penawaran AC LG - GD Checkup Laborat RS PHCДокумент1 страницаPenawaran AC LG - GD Checkup Laborat RS PHCaisyahbrillianaОценок пока нет
- Business Cognate SBAДокумент20 страницBusiness Cognate SBAUncle TravisОценок пока нет
- Affective DomainДокумент3 страницыAffective DomainJm Enriquez Dela Cruz50% (2)
- DataStage Interview QuestionsДокумент3 страницыDataStage Interview QuestionsvrkesariОценок пока нет
- Critical Period - Paper PsycholinguisticsДокумент6 страницCritical Period - Paper PsycholinguisticsDamas Dwi Rianto PutraОценок пока нет
- SWAY GuideДокумент16 страницSWAY Guide63456 touristaОценок пока нет
- Heat Transfer in Stagnation-Point Flow Towards A Stretching SheetДокумент5 страницHeat Transfer in Stagnation-Point Flow Towards A Stretching SheetboiroyОценок пока нет
- Document 1 PetДокумент9 страницDocument 1 PetRoel Jr Pinaroc DolaypanОценок пока нет
- Specification of SGP InterlayerДокумент3 страницыSpecification of SGP InterlayerHAN HANОценок пока нет
- Report Swtich Cisco Pass DNSДокумент91 страницаReport Swtich Cisco Pass DNSDenis Syst Laime LópezОценок пока нет
- Sop For RetailДокумент13 страницSop For Retailkarthika suresh100% (6)
- Learn C++ Programming LanguageДокумент322 страницыLearn C++ Programming LanguageAli Eb100% (8)
- Herzlich Willkommen - Welcome: German Information Session For New ParentsДокумент20 страницHerzlich Willkommen - Welcome: German Information Session For New ParentssammlissОценок пока нет
- PERFORM Toolkit 3 1 Release Notes 1.0 OnlinePDFДокумент18 страницPERFORM Toolkit 3 1 Release Notes 1.0 OnlinePDFJose HugoОценок пока нет
- Disneyland (Manish N Suraj)Документ27 страницDisneyland (Manish N Suraj)Suraj Kedia0% (1)
- WMS WIP PickДокумент14 страницWMS WIP PickAvinash RoutrayОценок пока нет
- Management - ControlДокумент20 страницManagement - Controlrsikira7905Оценок пока нет
- 27 EtdsДокумент29 страниц27 EtdsSuhag PatelОценок пока нет
- Do You Have Esp FinalДокумент20 страницDo You Have Esp FinalVrasidas Poulopoulos50% (2)
- Compiled H.W - Class 10th-1Документ3 страницыCompiled H.W - Class 10th-1Sree KalaОценок пока нет
- My Publications and Workshops ListДокумент4 страницыMy Publications and Workshops Listraviteja tankalaОценок пока нет
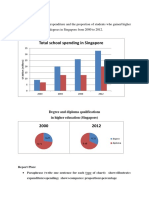
- Total School Spending in Singapore: Task 1 020219Документ6 страницTotal School Spending in Singapore: Task 1 020219viper_4554Оценок пока нет
- Vertical Axis Wind Turbine ProjДокумент2 страницыVertical Axis Wind Turbine Projmacsan sanchezОценок пока нет
- Practice Makes Perfect English Conversation UNIT 9Документ10 страницPractice Makes Perfect English Conversation UNIT 9Thaís C C MirandaОценок пока нет
- Culvert Design Write UpДокумент8 страницCulvert Design Write UpifylasyОценок пока нет
- Senior Project Manager Marketing in Toronto Canada Resume Cindy KempДокумент2 страницыSenior Project Manager Marketing in Toronto Canada Resume Cindy KempCindyKempОценок пока нет
- Morning Report: Supervisor: Dr. H. Doddy Ak., Spog (K)Документ10 страницMorning Report: Supervisor: Dr. H. Doddy Ak., Spog (K)vika handayaniОценок пока нет
- Complaint For Declaratory and Injunctive Relief FINALДокумент40 страницComplaint For Declaratory and Injunctive Relief FINALGilbert CordovaОценок пока нет