Вам также может понравиться
- Design of Cmos Transceiver For Micro-Nmr Spectrometer - Jaehyup KimДокумент147 страницDesign of Cmos Transceiver For Micro-Nmr Spectrometer - Jaehyup KimfergusoniseОценок пока нет
- Method For Determination of Transfer FunctionsДокумент5 страницMethod For Determination of Transfer Functionsjuandiegoc12Оценок пока нет
- 2011 - Powering Autonomous Sensors - PenellaДокумент155 страниц2011 - Powering Autonomous Sensors - PenellafergusoniseОценок пока нет
- Frequency Response Analysis and Design PDFДокумент281 страницаFrequency Response Analysis and Design PDFfergusoniseОценок пока нет
- Method For Determination of Transfer FunctionsДокумент5 страницMethod For Determination of Transfer Functionsjuandiegoc12Оценок пока нет
- Use of The Michigan Neuropathy Screening Instrument-2012Документ20 страницUse of The Michigan Neuropathy Screening Instrument-2012fergusoniseОценок пока нет
- Metabolic and Structural Role of Thiamine in Nervous-2008Документ9 страницMetabolic and Structural Role of Thiamine in Nervous-2008fergusoniseОценок пока нет
- Molecular Mechanisms of Thiamine Utilization: Charles K. Singleton and Peter R. MartinДокумент11 страницMolecular Mechanisms of Thiamine Utilization: Charles K. Singleton and Peter R. MartinfergusoniseОценок пока нет
- Perform Signal Integrity Analyses in Altium DesignerДокумент27 страницPerform Signal Integrity Analyses in Altium DesignerfergusoniseОценок пока нет
- Simple Mosfet ModelingДокумент8 страницSimple Mosfet ModelingfergusoniseОценок пока нет
- Least Squares ExampleДокумент2 страницыLeast Squares ExamplefergusoniseОценок пока нет
- FolkloreДокумент138 страницFolklorefergusoniseОценок пока нет
- Electronic SymbolsДокумент1 страницаElectronic SymbolsfergusoniseОценок пока нет
- Enc28j60 With Out WOLДокумент98 страницEnc28j60 With Out WOLManitОценок пока нет
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeОт EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeРейтинг: 4 из 5 звезд4/5 (5794)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceОт EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceРейтинг: 4 из 5 звезд4/5 (890)
- The Yellow House: A Memoir (2019 National Book Award Winner)От EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Рейтинг: 4 из 5 звезд4/5 (98)
- The Little Book of Hygge: Danish Secrets to Happy LivingОт EverandThe Little Book of Hygge: Danish Secrets to Happy LivingРейтинг: 3.5 из 5 звезд3.5/5 (399)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryОт EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryРейтинг: 3.5 из 5 звезд3.5/5 (231)
- Never Split the Difference: Negotiating As If Your Life Depended On ItОт EverandNever Split the Difference: Negotiating As If Your Life Depended On ItРейтинг: 4.5 из 5 звезд4.5/5 (838)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureОт EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureРейтинг: 4.5 из 5 звезд4.5/5 (474)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersОт EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersРейтинг: 4.5 из 5 звезд4.5/5 (344)
- The Emperor of All Maladies: A Biography of CancerОт EverandThe Emperor of All Maladies: A Biography of CancerРейтинг: 4.5 из 5 звезд4.5/5 (271)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaОт EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaРейтинг: 4.5 из 5 звезд4.5/5 (265)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreОт EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreРейтинг: 4 из 5 звезд4/5 (1090)
- Team of Rivals: The Political Genius of Abraham LincolnОт EverandTeam of Rivals: The Political Genius of Abraham LincolnРейтинг: 4.5 из 5 звезд4.5/5 (234)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyОт EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyРейтинг: 3.5 из 5 звезд3.5/5 (2219)
- The Unwinding: An Inner History of the New AmericaОт EverandThe Unwinding: An Inner History of the New AmericaРейтинг: 4 из 5 звезд4/5 (45)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)От EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Рейтинг: 4.5 из 5 звезд4.5/5 (119)
- Brain Respiratory Centers & Control MechanismsДокумент36 страницBrain Respiratory Centers & Control MechanismsJerilee SoCute WattsОценок пока нет
- Salbutamol Sulphate Inhaler for Asthma ReliefДокумент5 страницSalbutamol Sulphate Inhaler for Asthma ReliefRajib MandalОценок пока нет
- ZXZXZXДокумент8 страницZXZXZXharvey777Оценок пока нет
- English for Nursing: Affirmative and WH QuestionsДокумент24 страницыEnglish for Nursing: Affirmative and WH QuestionsLili FitrianiОценок пока нет
- PTSDДокумент9 страницPTSDAnisa Putri100% (1)
- HyponatremiaДокумент44 страницыHyponatremiaALi TaLib ShukurОценок пока нет
- Ancient Egypt's Views on Light, Blindness, and Eye HealthДокумент18 страницAncient Egypt's Views on Light, Blindness, and Eye HealthAdnan BuljubašićОценок пока нет
- ChronOS InjecДокумент24 страницыChronOS Injecmolecule112Оценок пока нет
- Usaf Nurse Corps-Information Booklet-4feb13Документ39 страницUsaf Nurse Corps-Information Booklet-4feb13Jorge Vigoreaux100% (1)
- Extrahepatic Biliary AtresiaДокумент3 страницыExtrahepatic Biliary AtresiaTiti Afrianto100% (1)
- Hands On Ivf Training - Chennai Fertility Centre - Chennai, India, DR - VM.ThomasДокумент21 страницаHands On Ivf Training - Chennai Fertility Centre - Chennai, India, DR - VM.ThomasThomasVadingaduОценок пока нет
- Pro MetricДокумент3 страницыPro Metricwandai_810% (1)
- Severe Ulcerative Colitis UHL Childrens Medical GuidelineДокумент8 страницSevere Ulcerative Colitis UHL Childrens Medical GuidelineFanny PritaningrumОценок пока нет
- 156.full (1) Badminton InjuriesДокумент5 страниц156.full (1) Badminton InjuriesRaul ValldecabresОценок пока нет
- Efinaconazole Topical Solution, 10%: Efficacy in Patients With Onychomycosis and Coexisting Tinea PedisДокумент5 страницEfinaconazole Topical Solution, 10%: Efficacy in Patients With Onychomycosis and Coexisting Tinea PedisTiara Rachmaputeri AriantoОценок пока нет
- A Survey of Stent Designs PDFДокумент11 страницA Survey of Stent Designs PDFGabriel Lopez BarajasОценок пока нет
- Muhammad Syauqi Mahamud Sayuti 071303051 Batch 21 Group E2Документ17 страницMuhammad Syauqi Mahamud Sayuti 071303051 Batch 21 Group E2Phong Qiu ShiОценок пока нет
- Naturopathic Treatment For Ear Pain in ChildrenДокумент8 страницNaturopathic Treatment For Ear Pain in ChildrenDr Siddharth SP YadavОценок пока нет
- Notification MUHS Teaching Non Teaching PostsДокумент2 страницыNotification MUHS Teaching Non Teaching PostsNeha SharmaОценок пока нет
- B.+BraunSpace 6064516 Pocket Guide PDFДокумент8 страницB.+BraunSpace 6064516 Pocket Guide PDFsusiloОценок пока нет
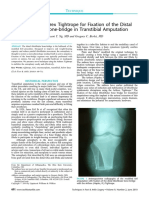
- Fixation of The Distal Tibiofibular Bone-Bridge in Transtibial Amputation PDFДокумент4 страницыFixation of The Distal Tibiofibular Bone-Bridge in Transtibial Amputation PDFThomas BowersОценок пока нет
- Josephine Ball ResumeДокумент1 страницаJosephine Ball Resumeapi-483700218Оценок пока нет
- PERINATAL PATIENT SAFETY: Management of Oxytocin For Labor Induction and Augmentation MCN, The American Journal of Maternal/Child NursingДокумент2 страницыPERINATAL PATIENT SAFETY: Management of Oxytocin For Labor Induction and Augmentation MCN, The American Journal of Maternal/Child NursingNishaThakuriОценок пока нет
- Regional Obstetric Early Warning Score ChartДокумент2 страницыRegional Obstetric Early Warning Score ChartDaeng Farahnaz100% (2)
- MAPO Index For Risk Assessment of Patient Manual Handling in Hospital Wards: A Validation StudyДокумент17 страницMAPO Index For Risk Assessment of Patient Manual Handling in Hospital Wards: A Validation StudySebastian LoperaОценок пока нет
- Residual Control ChartДокумент7 страницResidual Control ChartAgus HidayatullahОценок пока нет
- SW Provider Directory 2018 1ST QTR - CompressedДокумент20 страницSW Provider Directory 2018 1ST QTR - CompressedStayWell InsuranceОценок пока нет
- Guideline For Medical Internship ProgramДокумент41 страницаGuideline For Medical Internship ProgramMufaddal Madraswala50% (2)
- The IT Band Friction SyndromeДокумент3 страницыThe IT Band Friction Syndromecarmall15Оценок пока нет
- Taquiarritmias Supraventriculares NEJMДокумент11 страницTaquiarritmias Supraventriculares NEJMCamilo VidalОценок пока нет