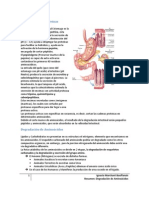
Вам также может понравиться
- Metabolismo de AminoácidosДокумент12 страницMetabolismo de AminoácidosGermán Sil100% (1)
- Ayudas ergogénicas y nutricionalesОт EverandAyudas ergogénicas y nutricionalesРейтинг: 5 из 5 звезд5/5 (2)
- Met. AaДокумент11 страницMet. AaPedro RodriguezОценок пока нет
- Resumen Proteinas y AminoacidosДокумент30 страницResumen Proteinas y Aminoacidosmarianne beltranОценок пока нет
- Proteolisis AminoacidosДокумент5 страницProteolisis AminoacidosLuis SantesОценок пока нет
- Funciones Clave de La Peptidil SerinaДокумент8 страницFunciones Clave de La Peptidil SerinaAG PCОценок пока нет
- HJHJJJKДокумент15 страницHJHJJJKSanti GarciaОценок пока нет
- Trabalho de Bioquimica 2Документ11 страницTrabalho de Bioquimica 2Thayssa De Souza Silva 3GОценок пока нет
- TextoДокумент4 страницыTextoAriana SilvaОценок пока нет
- Metabolismo de AminoacidosДокумент3 страницыMetabolismo de AminoacidosErikaDesimoniОценок пока нет
- Sem 12 File 3Документ26 страницSem 12 File 3Daniela MatamalaОценок пока нет
- AA GlucogénicosДокумент2 страницыAA GlucogénicosGabriela PrimorОценок пока нет
- Tema 18 Ruta de Los Aminoacidos. Autor Alexander Sanchez Lamas.Документ14 страницTema 18 Ruta de Los Aminoacidos. Autor Alexander Sanchez Lamas.Alexander SanchezОценок пока нет
- Resuemen de BQM Metabolismo y Degradación de AAДокумент2 страницыResuemen de BQM Metabolismo y Degradación de AAOldemar GarciaОценок пока нет
- Metabolismo de Los AminoácidosДокумент4 страницыMetabolismo de Los AminoácidosGuillermo Jose PerezОценок пока нет
- Resumen de Catabolismo de AminoácidosДокумент4 страницыResumen de Catabolismo de AminoácidosANTHONY CAMARGOОценок пока нет
- Metabolismo de Aminoácidos: Prof: Nerio SocorroДокумент47 страницMetabolismo de Aminoácidos: Prof: Nerio SocorroMildriana RiveroОценок пока нет
- Destinos Del Grupo AminoДокумент2 страницыDestinos Del Grupo AminoclaudiaОценок пока нет
- Metabolismo de AminoacidosДокумент3 страницыMetabolismo de AminoacidosAntonella BeratzОценок пока нет
- Aminoácidos. Equipo 2Документ2 страницыAminoácidos. Equipo 2FridaОценок пока нет
- Metabolismo Proteínas-EstДокумент32 страницыMetabolismo Proteínas-Estgerson1contreras-1Оценок пока нет
- Metabolismoi de AminiacidosДокумент7 страницMetabolismoi de Aminiacidosanon_609905353Оценок пока нет
- Resumen Degradación AminoacidosДокумент6 страницResumen Degradación AminoacidosIgnacio MarchantОценок пока нет
- Catabolismo de Los AminoácidosДокумент39 страницCatabolismo de Los AminoácidosEd BozaОценок пока нет
- RESUMEN. Metabolismo de Los Compuestos NitrogenadosДокумент8 страницRESUMEN. Metabolismo de Los Compuestos NitrogenadosFernanda AraizaОценок пока нет
- Tema 3 - Metabolismo de Aá y NucleótidosДокумент12 страницTema 3 - Metabolismo de Aá y NucleótidosepepОценок пока нет
- MetabolisДокумент10 страницMetabolisLuisa LopezОценок пока нет
- Degradación de Compuestos Nitrogenados.Документ19 страницDegradación de Compuestos Nitrogenados.Zulema Velázquez GarcíaОценок пока нет
- Metabolismo de Proteinas ResumenДокумент77 страницMetabolismo de Proteinas ResumenIris zavalaОценок пока нет
- Tema 3 - Metabolismo de A y Nucle TidosДокумент12 страницTema 3 - Metabolismo de A y Nucle TidosJON LEGUINAОценок пока нет
- Seminario Metabolismo de Aminoácidos 2Документ3 страницыSeminario Metabolismo de Aminoácidos 2Andrés SalazarОценок пока нет
- MetabolismoProteinas - Jacqueline VelascoДокумент6 страницMetabolismoProteinas - Jacqueline VelascoADRIANA ZUBIA FLORESОценок пока нет
- Seminario 10 Catabolismo de AminoácidosДокумент5 страницSeminario 10 Catabolismo de AminoácidosVictor CespedesОценок пока нет
- Introducción Al Metabolismo de Los AminoácidosДокумент27 страницIntroducción Al Metabolismo de Los AminoácidosmadeleynecyОценок пока нет
- Bio Qui MicaДокумент9 страницBio Qui MicaJatnielОценок пока нет
- Bioquimica II Primer ParcialДокумент48 страницBioquimica II Primer ParcialEmi MaciasОценок пока нет
- La Reacción de Glutamato DeshidrogenasaLa Reacción Catalizada Por La Glutamato Deshidrogenasa EsДокумент2 страницыLa Reacción de Glutamato DeshidrogenasaLa Reacción Catalizada Por La Glutamato Deshidrogenasa EsBrenda TorresОценок пока нет
- Desaminación Oxidativa ResumenДокумент2 страницыDesaminación Oxidativa ResumenChristopher Marte BaezОценок пока нет
- Utilización de Aminoácidos Absorbidos y Mecanismos de OxidaciónДокумент20 страницUtilización de Aminoácidos Absorbidos y Mecanismos de OxidaciónGenyta Cauich75% (4)
- 3 Metabolismo de Proteinas 2016Документ11 страниц3 Metabolismo de Proteinas 2016Estela EscalanteОценок пока нет
- Apunte Metabolismo de Los AminoácidosДокумент2 страницыApunte Metabolismo de Los AminoácidosCelma ZúñigaОценок пока нет
- Destino Del AminoДокумент6 страницDestino Del AminoJosé Alberto A MonteroОценок пока нет
- Desaminacion TransaminacionДокумент10 страницDesaminacion TransaminacionHector Manuel Lendo HernandezОценок пока нет
- Reacciones Metabólicas Generales de Los AminoácidosДокумент2 страницыReacciones Metabólicas Generales de Los AminoácidosJesus DavilaОценок пока нет
- Transporte de Amoniaco, Ciclo de La Urea y Catabolismo de Los Esqueletos CarbonadosДокумент5 страницTransporte de Amoniaco, Ciclo de La Urea y Catabolismo de Los Esqueletos CarbonadosMarina CarreonОценок пока нет
- Tema13 Catabolismo AminoacidosДокумент36 страницTema13 Catabolismo AminoacidosÁngela Díaz ChamorroОценок пока нет
- Catabolismo de AminoacidosДокумент20 страницCatabolismo de AminoacidosXimena Véliz DaudОценок пока нет
- Transaminación y Desaminación de ProteínasДокумент24 страницыTransaminación y Desaminación de ProteínasCITLALLI MU�OZ ANDRADEОценок пока нет
- Transaminaciones y Ciclo de La UreaДокумент16 страницTransaminaciones y Ciclo de La UreaRosario LagunaОценок пока нет
- Oxidación Aminoácidos PDFДокумент16 страницOxidación Aminoácidos PDFsilviaОценок пока нет
- Rutas PDFДокумент7 страницRutas PDFAdriana ProañoОценок пока нет
- Metabolismo de ProteinasДокумент20 страницMetabolismo de ProteinasYenny Orellana SolanoОценок пока нет
- Capítulo 11 Compuestos Nitrogenados No ProteicosДокумент13 страницCapítulo 11 Compuestos Nitrogenados No ProteicosDiego NolascoОценок пока нет
- Exposicion de Bioquimica 1Документ2 страницыExposicion de Bioquimica 1Giampierre VillenaОценок пока нет
- Metabolismo de Las ProteínasДокумент14 страницMetabolismo de Las ProteínasMaria Andrea CorredorОценок пока нет
- Metabolismo de Las ProteínasДокумент3 страницыMetabolismo de Las ProteínasKaren FernandaОценок пока нет
- Ciclo de La UreaДокумент5 страницCiclo de La Urealizesther2005Оценок пока нет
- 6Документ50 страниц6Oz HdzОценок пока нет
- Taller Metabolismo NitrógenoДокумент6 страницTaller Metabolismo NitrógenoLAURA FERNANDA HURTADO BERNALОценок пока нет
- Ocupación y PleurostomíaДокумент21 страницаOcupación y PleurostomíaDiego Fidelli LopezОценок пока нет
- Guía ClínicaДокумент363 страницыGuía ClínicaCésar Ibáñez MiñanoОценок пока нет
- Guía Pokemon Perla y DiamanteДокумент109 страницGuía Pokemon Perla y DiamanteDiego Fidelli LopezОценок пока нет
- Gran Quemado, Guia Minsal ChileДокумент49 страницGran Quemado, Guia Minsal Chiledrlarzouse100% (1)
- MonografíaДокумент23 страницыMonografíaDiego Fidelli LopezОценок пока нет
- Guia de EjerciciosixДокумент4 страницыGuia de EjerciciosixDiego Fidelli LopezОценок пока нет
- COMPARACIONДокумент1 страницаCOMPARACIONDiego Fidelli LopezОценок пока нет
- Declaracion Directivos 16 JunioДокумент2 страницыDeclaracion Directivos 16 JunioDiego Fidelli LopezОценок пока нет
- Introducción A LA Farmacognosia: VimcvfmДокумент30 страницIntroducción A LA Farmacognosia: VimcvfmIris Serrano VelasquezОценок пока нет
- Cuerpos CetonicosДокумент5 страницCuerpos CetonicosAngelesTorresОценок пока нет
- Leccion 1. SuelosДокумент17 страницLeccion 1. SuelosKatia Margarita Almanza BОценок пока нет
- Esquema BioquímicaДокумент2 страницыEsquema BioquímicaIsabel Maria Durán ZamoraОценок пока нет
- Metabolismo de Los Lipidos ResumenДокумент2 страницыMetabolismo de Los Lipidos ResumenPaco Malagon Bedoy100% (3)
- Enzimas DesnaturalizaciónДокумент16 страницEnzimas DesnaturalizaciónCristhian OlanoОценок пока нет
- 05 - Trabajo Practico #3 - Papa y Problemas - 2018 PDFДокумент6 страниц05 - Trabajo Practico #3 - Papa y Problemas - 2018 PDFCarli CastilloОценок пока нет
- Evaluación 7° Metabolismo y OsmorregulaciónДокумент1 страницаEvaluación 7° Metabolismo y OsmorregulaciónLuz Mary Betin100% (1)
- Carbohidratos y Proteinas-MonografiaДокумент59 страницCarbohidratos y Proteinas-MonografiaJhardy LucianoОценок пока нет
- Reacción HillДокумент8 страницReacción HillKimberly Torres100% (1)
- G Fisiopatologia HepaticaДокумент21 страницаG Fisiopatologia HepaticaMonaLocaОценок пока нет
- Hipoglicemia e HipocalcemiaДокумент31 страницаHipoglicemia e HipocalcemiaBRAYAN HANS PINEDO RODRIGUEZОценок пока нет
- Bioquimica IIДокумент6 страницBioquimica IIWilliam FernandoОценок пока нет
- Mapas CuadernillloДокумент52 страницыMapas Cuadernillloguisel peraltaОценок пока нет
- Compilacion Trabajo Colaborativo E INDIVIDUALДокумент58 страницCompilacion Trabajo Colaborativo E INDIVIDUALYessica Quimbayo AlvaresОценок пока нет
- Pruebas BioquimicasДокумент6 страницPruebas Bioquimicasarinayed_niwde4709Оценок пока нет
- Taller Metabolisma 7Документ2 страницыTaller Metabolisma 7Anonymous 1eW4s4Оценок пока нет
- Caso Clinico 2Документ2 страницыCaso Clinico 2Julio Estuado Orozco ChitíОценок пока нет
- 7 - Curso de FangoterapiaДокумент15 страниц7 - Curso de FangoterapiaGabriela SolariОценок пока нет
- Taller Sueroterapia MetabolicaДокумент14 страницTaller Sueroterapia MetabolicaKarol Andrade Quiñonez100% (2)
- Silabo Bioquimica 2015 II UNALMДокумент4 страницыSilabo Bioquimica 2015 II UNALMxmarioxxОценок пока нет
- Nuevo PPT GEOMETRIA SagradaДокумент36 страницNuevo PPT GEOMETRIA SagradabrujitasexyОценок пока нет
- Practica de Seres Vivos 2° Secundaria 2022Документ3 страницыPractica de Seres Vivos 2° Secundaria 2022Who is?Оценок пока нет
- VITAMINA1Документ2 страницыVITAMINA1Santos PalenciaОценок пока нет
- Diccionario Biología MolecularДокумент22 страницыDiccionario Biología MolecularHeffer Rueda EspinosaОценок пока нет
- Act ClaseДокумент16 страницAct ClaseAdrian Camilo MONROYОценок пока нет
- Carbohidratos BioquimicaДокумент4 страницыCarbohidratos BioquimicabeberlyОценок пока нет
- Insulina, Glucagon y AdrenalinaДокумент7 страницInsulina, Glucagon y AdrenalinaAlma EspinozaОценок пока нет
- Exámenes de Bioquímica PDFДокумент96 страницExámenes de Bioquímica PDFNicole MastrogiorgioОценок пока нет
- Práctica1 EspectrofotometríaДокумент6 страницPráctica1 EspectrofotometríaKaren AriasОценок пока нет