Вам также может понравиться
- Cell Press Reviews: Core Concepts in Cell BiologyОт EverandCell Press Reviews: Core Concepts in Cell BiologyCell PressОценок пока нет
- Cell Cycle and Cell DivisionДокумент8 страницCell Cycle and Cell Divisionbhupeshgalani100% (2)
- Bacterial Growth and Division: Biochemistry and Regulation of Prokaryotic and Eukaryotic Division CyclesОт EverandBacterial Growth and Division: Biochemistry and Regulation of Prokaryotic and Eukaryotic Division CyclesРейтинг: 5 из 5 звезд5/5 (1)
- Transport Across Cell MembraneДокумент38 страницTransport Across Cell MembraneViswadeep DasОценок пока нет
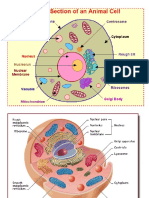
- CellДокумент32 страницыCellSrinivas Nakka100% (1)
- Cell TheoryДокумент10 страницCell TheoryLuigi Miguel G. Tirana100% (1)
- 2.4 - Membranes: 2.4.1 - Draw and Label A Diagram To Show The Structure of MembranesДокумент6 страниц2.4 - Membranes: 2.4.1 - Draw and Label A Diagram To Show The Structure of MembranesDuong TongОценок пока нет
- Advanced Biology: PhotosynthesisДокумент31 страницаAdvanced Biology: Photosynthesiszorbax100% (2)
- Cellular and Molecular Mechanisms of Inflammation: Receptors of Inflammatory Cells: Structure—Function RelationshipsОт EverandCellular and Molecular Mechanisms of Inflammation: Receptors of Inflammatory Cells: Structure—Function RelationshipsCharles G. CochraneОценок пока нет
- 2.4.1 Cell WallДокумент17 страниц2.4.1 Cell WallSyahirah SyerryОценок пока нет
- Transport Across Membranes FinalДокумент77 страницTransport Across Membranes FinalLorraine Eve Abila100% (1)
- HDTD-B-4 - Cell OrganellesДокумент43 страницыHDTD-B-4 - Cell OrganellesMariam Qais100% (1)
- Plasma MembraneДокумент35 страницPlasma MembraneMini RůžičkaОценок пока нет
- Cellular Respiration PDFДокумент10 страницCellular Respiration PDFAnonymous HXLczq3Оценок пока нет
- Cell Growth and DivisionДокумент15 страницCell Growth and DivisionJohn Lester BerdinОценок пока нет
- Transport in PlantsДокумент21 страницаTransport in PlantsMothi KarunaОценок пока нет
- Cell Structure and FunctionДокумент41 страницаCell Structure and FunctionAhkyluzLaniaz100% (1)
- Cell Structure & FunctionДокумент39 страницCell Structure & Functiondacspinlac100% (9)
- Cell Membrane TransportДокумент75 страницCell Membrane Transportvinay chaudhary100% (1)
- Cell DivisionДокумент70 страницCell Divisioncandice swanepoel100% (2)
- CellsДокумент24 страницыCellsapi-95529788100% (1)
- 1..cell Structure and FunctionДокумент53 страницы1..cell Structure and FunctionAminath MeesanОценок пока нет
- Ch5 Cell MembranesДокумент60 страницCh5 Cell MembranesAdrian Aguilera GarciaОценок пока нет
- Chapter 6: Cell Division: 6.2: Cell Cycle and MitosisДокумент18 страницChapter 6: Cell Division: 6.2: Cell Cycle and MitosisMohammed Ridzuwan0% (1)
- p3 - Cellular Transport MechanismДокумент53 страницыp3 - Cellular Transport MechanismJudithОценок пока нет
- Chloroplast: The Structural Site of PhotosynthesisДокумент4 страницыChloroplast: The Structural Site of PhotosynthesisNathaliaEicellRoseBuenoОценок пока нет
- Cel Mol LecДокумент3 страницыCel Mol LecIzza LimОценок пока нет
- CH 11 Cell CommunicationДокумент4 страницыCH 11 Cell Communicationwil7ver100% (2)
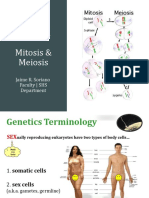
- Mitosis and MeiosisДокумент30 страницMitosis and MeiosisAmalina Ismail100% (4)
- Enzymes WorksheetДокумент8 страницEnzymes WorksheetClaudia PinnaОценок пока нет
- Introduction To EnzymesДокумент43 страницыIntroduction To EnzymesVytheeshwaran Vedagiri100% (2)
- General Biology Mid Exam Summary Semester OneДокумент26 страницGeneral Biology Mid Exam Summary Semester OneDeborah ShirleenОценок пока нет
- Mitosis Meiosis NOTESДокумент11 страницMitosis Meiosis NOTESCaden DiazОценок пока нет
- The Cell Cycle Worksheet With AnswersДокумент3 страницыThe Cell Cycle Worksheet With AnswersAngelic ZapantaОценок пока нет
- Cell MembraneДокумент100 страницCell MembranephyMedza AppОценок пока нет
- Prokaryotic and Eukaryotic CellsДокумент18 страницProkaryotic and Eukaryotic Cellssalahuddin_md5935100% (1)
- Chapter 5 Cell DivisionДокумент18 страницChapter 5 Cell DivisionEma Fatimah0% (1)
- Cell Biology LaboratoryДокумент319 страницCell Biology LaboratoryNazaqat FarooqОценок пока нет
- Cell Membrane Coloring Worksheet - KEYДокумент6 страницCell Membrane Coloring Worksheet - KEYsapphire284583% (12)
- Cell Cycle (Cell Division)Документ60 страницCell Cycle (Cell Division)Janardhan Reddy P V100% (2)
- Mitosis and MeiosisДокумент56 страницMitosis and MeiosisJaime Soriano100% (2)
- Cell Membrane & Tonicity WorksheetДокумент5 страницCell Membrane & Tonicity WorksheetSofia Amelia SuryaniОценок пока нет
- CellДокумент15 страницCellprakash kushwahaОценок пока нет
- WORKSHEET 5.7 Comparing Mitosis and MeiosisДокумент4 страницыWORKSHEET 5.7 Comparing Mitosis and Meiosiswienna1987Оценок пока нет
- Cell Membranes and TransportДокумент10 страницCell Membranes and Transportlina100% (2)
- Chloroplast NotesДокумент4 страницыChloroplast NotesAnanda100% (1)
- General Biology 1: The CellsДокумент71 страницаGeneral Biology 1: The CellsPaula Marie Llido100% (2)
- Oxidative PhosphorylationДокумент33 страницыOxidative PhosphorylationJithendra Babu0% (1)
- Regulation of Cell CycleДокумент5 страницRegulation of Cell CyclemohsinОценок пока нет
- Overview of Cell Cycle by Javali.GДокумент15 страницOverview of Cell Cycle by Javali.GJavali.GОценок пока нет
- Cell Biology: The Cell Division CylceДокумент56 страницCell Biology: The Cell Division Cylcebabybri94Оценок пока нет
- Q1 Module 1 The Cell TheoryДокумент29 страницQ1 Module 1 The Cell TheoryKathy BaynosaОценок пока нет
- Cell and Cell TheoryДокумент13 страницCell and Cell TheoryAnaJeanJumawidMaranan100% (1)
- Cell Parts and Its FunctionsДокумент25 страницCell Parts and Its FunctionsSen Armario100% (1)
- Gel ElectrophoresisДокумент6 страницGel Electrophoresisapi-385481351Оценок пока нет
- Intermediate FilamentsДокумент4 страницыIntermediate FilamentsSai SridharОценок пока нет
- Digital Communications I: Modulation and Coding Course: Term 3 - 2008 Catharina LogothetisДокумент27 страницDigital Communications I: Modulation and Coding Course: Term 3 - 2008 Catharina LogothetiserichaasОценок пока нет
- Digital Communication I: Modulation and Coding Course: Term 3 - 2009Документ28 страницDigital Communication I: Modulation and Coding Course: Term 3 - 2009erichaasОценок пока нет
- Digital Communications I: Modulation and Coding Course: Period 3 - 2007 Catharina LogothetisДокумент30 страницDigital Communications I: Modulation and Coding Course: Period 3 - 2007 Catharina LogothetiserichaasОценок пока нет
- TL Tus MFL39760283 17 191202 00 WebДокумент157 страницTL Tus MFL39760283 17 191202 00 WeberichaasОценок пока нет
- Digital Communications I: Modulation and Coding Course: Term 3 - 2008 Catharina LogothetisДокумент25 страницDigital Communications I: Modulation and Coding Course: Term 3 - 2008 Catharina LogothetiserichaasОценок пока нет
- Digital Communications I: Modulation and Coding Course: Term 3 - 2008 Catharina LogothetisДокумент21 страницаDigital Communications I: Modulation and Coding Course: Term 3 - 2008 Catharina LogothetiserichaasОценок пока нет
- Face Mask Pattern: 4 Sizes, Plus A Video TutorialДокумент6 страницFace Mask Pattern: 4 Sizes, Plus A Video TutorialerichaasОценок пока нет
- Multirate Signal ProcessingДокумент54 страницыMultirate Signal ProcessingerichaasОценок пока нет
- Digital Communications I: Modulation and Coding Course: Term 3 - 2008 Catharina LogothetisДокумент22 страницыDigital Communications I: Modulation and Coding Course: Term 3 - 2008 Catharina LogothetiserichaasОценок пока нет
- Chap 00 IntroДокумент11 страницChap 00 IntroerichaasОценок пока нет
- GR1268 SolutionsДокумент46 страницGR1268 SolutionserichaasОценок пока нет
- CH 38 Cellular MotilityДокумент19 страницCH 38 Cellular MotilityerichaasОценок пока нет
- How To Play Craps - 11 Rolls To WinДокумент16 страницHow To Play Craps - 11 Rolls To WinerichaasОценок пока нет
- CH - 44 - Mitosis and Cytokinesis PDFДокумент24 страницыCH - 44 - Mitosis and Cytokinesis PDFerichaasОценок пока нет
- CH 36 Motor ProteinsДокумент16 страницCH 36 Motor ProteinserichaasОценок пока нет
- Nawcwd TP 8347 PDFДокумент455 страницNawcwd TP 8347 PDFerichaasОценок пока нет
- CH - 37 - Intracellular Motility PDFДокумент12 страницCH - 37 - Intracellular Motility PDFerichaasОценок пока нет
- CH 10 Gene ExpressionДокумент23 страницыCH 10 Gene ExpressionerichaasОценок пока нет
- CH 39 MusclesДокумент21 страницаCH 39 MuscleserichaasОценок пока нет
- How A Math Nerd Plays CrapsДокумент75 страницHow A Math Nerd Plays Crapserichaas100% (4)
- Illumina Bead ChipsДокумент5 страницIllumina Bead ChipsJuan RamirezОценок пока нет
- UQ ST Lucia MapДокумент1 страницаUQ ST Lucia MapqusoiriОценок пока нет
- Futura Primus Flexible Great Toe FinalДокумент12 страницFutura Primus Flexible Great Toe FinalNasfikurОценок пока нет
- PharmaДокумент11 страницPharmaRai SenguptaОценок пока нет
- International Journal On Bioinformatics & Biosciences (IJBB)Документ1 страницаInternational Journal On Bioinformatics & Biosciences (IJBB)Anonymous pKuPK3zUОценок пока нет
- History of GeneticsДокумент4 страницыHistory of GeneticsJanine FandaganiОценок пока нет
- RANKL Is Associated With Persistent Primary Teeth and Delayed Permanent Tooth EmergenceДокумент8 страницRANKL Is Associated With Persistent Primary Teeth and Delayed Permanent Tooth EmergencefghdhmdkhОценок пока нет
- Significance of The Patched Gene in Developmental Biology: Evan Zhou T05 CMMB 403Документ9 страницSignificance of The Patched Gene in Developmental Biology: Evan Zhou T05 CMMB 403EvanОценок пока нет
- New Microsoft Word DocumentДокумент4 страницыNew Microsoft Word DocumentGina Portuguese GawonОценок пока нет
- Dot Blot HandoutДокумент11 страницDot Blot HandoutLeana KireiОценок пока нет
- Practice Questions For Dna Replication and RepairДокумент11 страницPractice Questions For Dna Replication and RepairdhruviniОценок пока нет
- Appl. Environ. Microbiol. 2011 Meyer AEM.05434 11Документ37 страницAppl. Environ. Microbiol. 2011 Meyer AEM.05434 11mrtharamОценок пока нет
- Iq Sybr Green Supermix: Instruction ManualДокумент20 страницIq Sybr Green Supermix: Instruction Manualkromatin9462Оценок пока нет
- Alexander Shekhtman, David S. Burz-Protein NMR Techniques (Methods in Molecular Biology, V831) - Springer (2011)Документ533 страницыAlexander Shekhtman, David S. Burz-Protein NMR Techniques (Methods in Molecular Biology, V831) - Springer (2011)Alan Raphael MoraesОценок пока нет
- Computational Biology, Part 8: Protein Coding RegionsДокумент40 страницComputational Biology, Part 8: Protein Coding RegionsDev AshwaniОценок пока нет
- Bioinformatic Tools and Guideline For PCR Primer DesignДокумент5 страницBioinformatic Tools and Guideline For PCR Primer DesignDeepak Ranjan SahooОценок пока нет
- Sartorius Bio Aseptic SamplingДокумент9 страницSartorius Bio Aseptic SamplingTimОценок пока нет
- 0.59062000 - 1469511231 - Bio Medical Waste Management SOP - 2Документ80 страниц0.59062000 - 1469511231 - Bio Medical Waste Management SOP - 2Alan MudaryОценок пока нет
- AST Testing MethodsДокумент11 страницAST Testing Methodstanty_ukОценок пока нет
- Lipofectamine ManualДокумент4 страницыLipofectamine ManualBui Ngoc ThuyОценок пока нет
- Use of Biotechnology in Agriculture-Benefits and RisksДокумент6 страницUse of Biotechnology in Agriculture-Benefits and RisksmortezahОценок пока нет
- A User's Guide To The Human Genome (2002)Документ82 страницыA User's Guide To The Human Genome (2002)Ilker BüyükОценок пока нет
- Heavy Metal Detection Using BioluminescenceДокумент26 страницHeavy Metal Detection Using BioluminescenceKaustubh ParekhОценок пока нет
- Wizard ProtocoloДокумент4 страницыWizard ProtocoloHarold Muñoz CiezaОценок пока нет
- MMS Project Guidelines RevisedДокумент73 страницыMMS Project Guidelines RevisedsilpaveeОценок пока нет
- List of Approved Journals: All Foreign Journals and Following Indian JournalsДокумент8 страницList of Approved Journals: All Foreign Journals and Following Indian JournalsAmit MishraОценок пока нет
- Mla Format Gmo PaperДокумент4 страницыMla Format Gmo Paperapi-279662666Оценок пока нет
- Biocon - HRDДокумент29 страницBiocon - HRDsreeramk13100% (1)
- Effect of Environmental Factors On The Growth of MicroorganismsДокумент10 страницEffect of Environmental Factors On The Growth of MicroorganismsHan Hui ChongОценок пока нет
- Bukti Monitoring InspeksiДокумент115 страницBukti Monitoring Inspeksielah murianiОценок пока нет
- All That Remains: A Renowned Forensic Scientist on Death, Mortality, and Solving CrimesОт EverandAll That Remains: A Renowned Forensic Scientist on Death, Mortality, and Solving CrimesРейтинг: 4.5 из 5 звезд4.5/5 (397)
- Gut: the new and revised Sunday Times bestsellerОт EverandGut: the new and revised Sunday Times bestsellerРейтинг: 4 из 5 звезд4/5 (393)
- Why We Die: The New Science of Aging and the Quest for ImmortalityОт EverandWhy We Die: The New Science of Aging and the Quest for ImmortalityРейтинг: 4 из 5 звезд4/5 (5)
- The Molecule of More: How a Single Chemical in Your Brain Drives Love, Sex, and Creativity--and Will Determine the Fate of the Human RaceОт EverandThe Molecule of More: How a Single Chemical in Your Brain Drives Love, Sex, and Creativity--and Will Determine the Fate of the Human RaceРейтинг: 4.5 из 5 звезд4.5/5 (517)
- When the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisОт EverandWhen the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisРейтинг: 3.5 из 5 звезд3.5/5 (2)
- 10% Human: How Your Body's Microbes Hold the Key to Health and HappinessОт Everand10% Human: How Your Body's Microbes Hold the Key to Health and HappinessРейтинг: 4 из 5 звезд4/5 (33)
- Tales from Both Sides of the Brain: A Life in NeuroscienceОт EverandTales from Both Sides of the Brain: A Life in NeuroscienceРейтинг: 3 из 5 звезд3/5 (18)
- A Brief History of Intelligence: Evolution, AI, and the Five Breakthroughs That Made Our BrainsОт EverandA Brief History of Intelligence: Evolution, AI, and the Five Breakthroughs That Made Our BrainsРейтинг: 4.5 из 5 звезд4.5/5 (6)
- The Ancestor's Tale: A Pilgrimage to the Dawn of EvolutionОт EverandThe Ancestor's Tale: A Pilgrimage to the Dawn of EvolutionРейтинг: 4 из 5 звезд4/5 (812)
- Return of the God Hypothesis: Three Scientific Discoveries That Reveal the Mind Behind the UniverseОт EverandReturn of the God Hypothesis: Three Scientific Discoveries That Reveal the Mind Behind the UniverseРейтинг: 4.5 из 5 звезд4.5/5 (52)
- Fast Asleep: Improve Brain Function, Lose Weight, Boost Your Mood, Reduce Stress, and Become a Better SleeperОт EverandFast Asleep: Improve Brain Function, Lose Weight, Boost Your Mood, Reduce Stress, and Become a Better SleeperРейтинг: 4.5 из 5 звезд4.5/5 (16)
- Who's in Charge?: Free Will and the Science of the BrainОт EverandWho's in Charge?: Free Will and the Science of the BrainРейтинг: 4 из 5 звезд4/5 (65)
- Gut: The Inside Story of Our Body's Most Underrated Organ (Revised Edition)От EverandGut: The Inside Story of Our Body's Most Underrated Organ (Revised Edition)Рейтинг: 4 из 5 звезд4/5 (411)
- Undeniable: How Biology Confirms Our Intuition That Life Is DesignedОт EverandUndeniable: How Biology Confirms Our Intuition That Life Is DesignedРейтинг: 4 из 5 звезд4/5 (11)
- Buddha's Brain: The Practical Neuroscience of Happiness, Love & WisdomОт EverandBuddha's Brain: The Practical Neuroscience of Happiness, Love & WisdomРейтинг: 4 из 5 звезд4/5 (216)
- Good Without God: What a Billion Nonreligious People Do BelieveОт EverandGood Without God: What a Billion Nonreligious People Do BelieveРейтинг: 4 из 5 звезд4/5 (66)
- The Other Side of Normal: How Biology Is Providing the Clues to Unlock the Secrets of Normal and Abnormal BehaviorОт EverandThe Other Side of Normal: How Biology Is Providing the Clues to Unlock the Secrets of Normal and Abnormal BehaviorОценок пока нет
- Seven and a Half Lessons About the BrainОт EverandSeven and a Half Lessons About the BrainРейтинг: 4 из 5 звезд4/5 (110)
- Human: The Science Behind What Makes Your Brain UniqueОт EverandHuman: The Science Behind What Makes Your Brain UniqueРейтинг: 3.5 из 5 звезд3.5/5 (38)
- Moral Tribes: Emotion, Reason, and the Gap Between Us and ThemОт EverandMoral Tribes: Emotion, Reason, and the Gap Between Us and ThemРейтинг: 4.5 из 5 звезд4.5/5 (115)
- The Rise and Fall of the Dinosaurs: A New History of a Lost WorldОт EverandThe Rise and Fall of the Dinosaurs: A New History of a Lost WorldРейтинг: 4 из 5 звезд4/5 (595)
- Minds Make Societies: How Cognition Explains the World Humans CreateОт EverandMinds Make Societies: How Cognition Explains the World Humans CreateРейтинг: 4.5 из 5 звезд4.5/5 (24)
- A Series of Fortunate Events: Chance and the Making of the Planet, Life, and YouОт EverandA Series of Fortunate Events: Chance and the Making of the Planet, Life, and YouРейтинг: 4.5 из 5 звезд4.5/5 (62)
- The Invention of Tomorrow: A Natural History of ForesightОт EverandThe Invention of Tomorrow: A Natural History of ForesightРейтинг: 4.5 из 5 звезд4.5/5 (5)
- The Lives of Bees: The Untold Story of the Honey Bee in the WildОт EverandThe Lives of Bees: The Untold Story of the Honey Bee in the WildРейтинг: 4.5 из 5 звезд4.5/5 (44)