Вам также может понравиться
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeОт EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeРейтинг: 4 из 5 звезд4/5 (5794)
- The Little Book of Hygge: Danish Secrets to Happy LivingОт EverandThe Little Book of Hygge: Danish Secrets to Happy LivingРейтинг: 3.5 из 5 звезд3.5/5 (400)
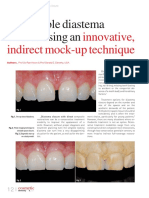
- Innovative, Indirect Mock-Up Technique: Predictable Diastema Closure Using AnДокумент4 страницыInnovative, Indirect Mock-Up Technique: Predictable Diastema Closure Using AnJing XueОценок пока нет
- 10.2319 080619-516EPubДокумент8 страниц10.2319 080619-516EPubJing XueОценок пока нет
- Case Report: Diastema Closure in Anterior Teeth Using A Posterior MatrixДокумент7 страницCase Report: Diastema Closure in Anterior Teeth Using A Posterior MatrixJing XueОценок пока нет
- Pak Orthod J 2013 5 1 27 33 PDFДокумент7 страницPak Orthod J 2013 5 1 27 33 PDFrief_candidaОценок пока нет
- ContentServer PDFДокумент7 страницContentServer PDFArchidHananAfifahОценок пока нет
- 10 1111@jopr 13109Документ6 страниц10 1111@jopr 13109Jing XueОценок пока нет
- 33-Article Text-71-1-10-20120907Документ5 страниц33-Article Text-71-1-10-20120907Jing XueОценок пока нет
- 10-Pramod Philip (Short Communication)Документ3 страницы10-Pramod Philip (Short Communication)Jing XueОценок пока нет
- 5-BarrosdeCamposetal 2015IJEDДокумент12 страниц5-BarrosdeCamposetal 2015IJEDJing XueОценок пока нет
- Closure of Diastema and Gingival Recontouring Using Direct Adhesive Restorations: A Case ReportДокумент12 страницClosure of Diastema and Gingival Recontouring Using Direct Adhesive Restorations: A Case ReportJing XueОценок пока нет
- Conservative Approach For The Esthetic Management of Multiple Interdental Spaces: A Systematic ApproachДокумент11 страницConservative Approach For The Esthetic Management of Multiple Interdental Spaces: A Systematic ApproachJing XueОценок пока нет
- 10 1111@jopr 13109Документ6 страниц10 1111@jopr 13109Jing XueОценок пока нет
- Midline Diastema Closure With Partial Laminate Veneers: A Case ReportДокумент4 страницыMidline Diastema Closure With Partial Laminate Veneers: A Case ReportAndykaYayanSetiawanОценок пока нет
- 0003-3219 (1977) 047 0265 Tmidai 2 0 Co 2Документ7 страниц0003-3219 (1977) 047 0265 Tmidai 2 0 Co 2Jing XueОценок пока нет
- Factors Affecting HealingДокумент12 страницFactors Affecting HealingpriyadharshiniОценок пока нет
- Survival of Restored Endodontically Treated Teeth in Relation To Periodontal StatusДокумент4 страницыSurvival of Restored Endodontically Treated Teeth in Relation To Periodontal StatusJing XueОценок пока нет
- Intraorifice Sealing Ability of Different Materials in Endodontically Treated TeethДокумент5 страницIntraorifice Sealing Ability of Different Materials in Endodontically Treated TeethJing XueОценок пока нет
- Midline Diastema Closure With Partial Laminate Veneers: A Case ReportДокумент4 страницыMidline Diastema Closure With Partial Laminate Veneers: A Case ReportAndykaYayanSetiawanОценок пока нет
- Incidence of Pain Associated With Clinical Factors During and After Root Canal Therapy. Part 1. Interappointment PainДокумент4 страницыIncidence of Pain Associated With Clinical Factors During and After Root Canal Therapy. Part 1. Interappointment PainJing XueОценок пока нет
- Do We Really Know How To Evaluate Tooth Prognosis A Systematic Review and Suggested Approach.1 PDFДокумент11 страницDo We Really Know How To Evaluate Tooth Prognosis A Systematic Review and Suggested Approach.1 PDFJing XueОценок пока нет
- Wide Diameter Implants-Degidi Piattelli LezziДокумент7 страницWide Diameter Implants-Degidi Piattelli LezziJing XueОценок пока нет
- Outcome of Root Canal Treatment in Dogs Determined by Periapical Radiography and Cone-Beam Computed Tomography ScansДокумент4 страницыOutcome of Root Canal Treatment in Dogs Determined by Periapical Radiography and Cone-Beam Computed Tomography ScansJing XueОценок пока нет
- Alve Olar Changes in Sockets Post ExtractionДокумент11 страницAlve Olar Changes in Sockets Post ExtractionPratyusha VallamОценок пока нет
- Dental Vertical Root Fractures: Value of CT in Detection: Head and Neck ImagingДокумент5 страницDental Vertical Root Fractures: Value of CT in Detection: Head and Neck ImagingJing XueОценок пока нет
- When To Refer To Specialist3Документ2 страницыWhen To Refer To Specialist3Jing XueОценок пока нет
- Walton SinusTractTracingEffectivenessДокумент1 страницаWalton SinusTractTracingEffectivenessJing XueОценок пока нет
- When To Refer To Specialist2Документ2 страницыWhen To Refer To Specialist2Jing XueОценок пока нет
- Why Clinicians Are Natural Bayesians: Interpreting Diagnostic Test Results From The Bayesian PerspectiveДокумент4 страницыWhy Clinicians Are Natural Bayesians: Interpreting Diagnostic Test Results From The Bayesian PerspectiveJing XueОценок пока нет
- Project Deliverable: Safety and Efficacy of A New and Emerging Dental X-Ray ModalityДокумент16 страницProject Deliverable: Safety and Efficacy of A New and Emerging Dental X-Ray ModalityJing XueОценок пока нет
- Avulsions and Intrusions: The Controversial Displacement InjuriesДокумент7 страницAvulsions and Intrusions: The Controversial Displacement InjuriesJing XueОценок пока нет
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceОт EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceРейтинг: 4 из 5 звезд4/5 (895)
- The Yellow House: A Memoir (2019 National Book Award Winner)От EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Рейтинг: 4 из 5 звезд4/5 (98)
- The Emperor of All Maladies: A Biography of CancerОт EverandThe Emperor of All Maladies: A Biography of CancerРейтинг: 4.5 из 5 звезд4.5/5 (271)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryОт EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryРейтинг: 3.5 из 5 звезд3.5/5 (231)
- Never Split the Difference: Negotiating As If Your Life Depended On ItОт EverandNever Split the Difference: Negotiating As If Your Life Depended On ItРейтинг: 4.5 из 5 звезд4.5/5 (838)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureОт EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureРейтинг: 4.5 из 5 звезд4.5/5 (474)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaОт EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaРейтинг: 4.5 из 5 звезд4.5/5 (266)
- The Unwinding: An Inner History of the New AmericaОт EverandThe Unwinding: An Inner History of the New AmericaРейтинг: 4 из 5 звезд4/5 (45)
- Team of Rivals: The Political Genius of Abraham LincolnОт EverandTeam of Rivals: The Political Genius of Abraham LincolnРейтинг: 4.5 из 5 звезд4.5/5 (234)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyОт EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyРейтинг: 3.5 из 5 звезд3.5/5 (2259)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreОт EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreРейтинг: 4 из 5 звезд4/5 (1090)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersОт EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersРейтинг: 4.5 из 5 звезд4.5/5 (344)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)От EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Рейтинг: 4.5 из 5 звезд4.5/5 (121)
- Hospital Manegment SoftwareДокумент6 страницHospital Manegment SoftwareShivpeth comОценок пока нет
- Heat StrokeДокумент4 страницыHeat StrokeGerald YasonОценок пока нет
- Docifu30002940j Perfecttcsii Ifu Eu Languagessallaindv1Документ196 страницDocifu30002940j Perfecttcsii Ifu Eu Languagessallaindv1Miguel De Los Santos PavisicОценок пока нет
- BHS InggrisДокумент4 страницыBHS InggrisarifinilhamОценок пока нет
- Lions Club Membership Application 111Документ2 страницыLions Club Membership Application 111Bolina, Pauline N.Оценок пока нет
- Stress MGMT in Call CentersДокумент70 страницStress MGMT in Call CentersJayesh Kambli100% (1)
- Looking Forward: Immigrant Contributions To The Golden State (2014)Документ2 страницыLooking Forward: Immigrant Contributions To The Golden State (2014)CALimmigrantОценок пока нет
- Accredited BAI Laboratory Animal Shelters 2018Документ8 страницAccredited BAI Laboratory Animal Shelters 2018ShanaiaОценок пока нет
- Course Unit Task: Teaching PlanДокумент2 страницыCourse Unit Task: Teaching PlanQueen100% (1)
- Useful BooksДокумент3 страницыUseful BooksphuongfeoОценок пока нет
- Study Plan FinalДокумент5 страницStudy Plan FinalMd IslamОценок пока нет
- Errors of Eye Refraction and HomoeopathyДокумент11 страницErrors of Eye Refraction and HomoeopathyDr. Rajneesh Kumar Sharma MD HomОценок пока нет
- (ThichTiengAnh - Com) de Theo Cau Truc Moi Nam 2017 de 1 Khong Dap AnДокумент101 страница(ThichTiengAnh - Com) de Theo Cau Truc Moi Nam 2017 de 1 Khong Dap AnRoland VietnamОценок пока нет
- Soares - Como Escolher Um Anestesico Local - 2005 PDFДокумент9 страницSoares - Como Escolher Um Anestesico Local - 2005 PDFjvb sobralОценок пока нет
- SOP For Cleaning of Ultrasound Probes in Trust Imaging Departments PDFДокумент2 страницыSOP For Cleaning of Ultrasound Probes in Trust Imaging Departments PDFShinta Vivoy21Оценок пока нет
- Steam Inhalation Definition:-: A. by Jug MethodДокумент4 страницыSteam Inhalation Definition:-: A. by Jug Methodss100% (1)
- Patient Transportation ProtocolДокумент8 страницPatient Transportation Protocolsami ketemaОценок пока нет
- STCW Section A-Vi.1Документ6 страницSTCW Section A-Vi.1Ernesto FariasОценок пока нет
- Block Learning Guide (BLG) : Block II Hematoimmunology System (HIS)Документ6 страницBlock Learning Guide (BLG) : Block II Hematoimmunology System (HIS)ASTAGINA NAURAHОценок пока нет
- Injuries Diseases & Disorders of The Muscular SystemДокумент22 страницыInjuries Diseases & Disorders of The Muscular SystemAngeli LozanoОценок пока нет
- S9 Q4 M1 Transport and Excretion and Respiration and Energy Exchange QuizДокумент3 страницыS9 Q4 M1 Transport and Excretion and Respiration and Energy Exchange QuizJermae DizonОценок пока нет
- How The Red Cross Raised Half A Billion Dollars For Haiti - and Built Six Homes - ProPublicaДокумент15 страницHow The Red Cross Raised Half A Billion Dollars For Haiti - and Built Six Homes - ProPublicaGina PistolОценок пока нет
- 017-2013 - MR (MOA Kalahi Cidss)Документ3 страницы017-2013 - MR (MOA Kalahi Cidss)SbGuinobatanОценок пока нет
- 1 SMДокумент14 страниц1 SMRosalina Pertiwi GultomОценок пока нет
- Amber Sewell: Professional SummaryДокумент4 страницыAmber Sewell: Professional Summaryapi-383979726Оценок пока нет
- Child Abuse Nursing Care PlanДокумент7 страницChild Abuse Nursing Care PlanMAHESH KOUJALAGIОценок пока нет
- The Demand For, and Impact Of, Learning HIV StatusДокумент36 страницThe Demand For, and Impact Of, Learning HIV StatusWilson LancherosОценок пока нет
- Nursing Role(s) in Emergency and DisasterДокумент44 страницыNursing Role(s) in Emergency and DisasterOEDIEN-kmbОценок пока нет
- HIIT Academy Total Body Blast Phase 1 PDFДокумент33 страницыHIIT Academy Total Body Blast Phase 1 PDFAndre ChinassoОценок пока нет
- Malaria Case AnalysisДокумент1 страницаMalaria Case AnalysisANGELA MARIE VILLENAОценок пока нет