Вам также может понравиться
- REVIEW - The Art of Fin Regeneration in Zebrafish - Catherine Pfefferli & Anna Jazwinska (2015)Документ12 страницREVIEW - The Art of Fin Regeneration in Zebrafish - Catherine Pfefferli & Anna Jazwinska (2015)Bagas Lantip PrakasaОценок пока нет
- Jonsson 2014Документ10 страницJonsson 2014Arvanza RivaieОценок пока нет
- Detailed Reconstruction of The Musculature in Limnognathia Maerski (Micrognathozoa) and Comparison With Other GnathiferaДокумент17 страницDetailed Reconstruction of The Musculature in Limnognathia Maerski (Micrognathozoa) and Comparison With Other GnathiferaDayana FlorezОценок пока нет
- Washington Academy of Sciences Journal of The Washington Academy of SciencesДокумент18 страницWashington Academy of Sciences Journal of The Washington Academy of SciencesGuillermo Alonso PovedaОценок пока нет
- Hörnschemeyer-2013 13 1-14Документ14 страницHörnschemeyer-2013 13 1-14Thomas HörnschemeyerОценок пока нет
- 3-The Newest Synthesis - Understanding The Interplay of Evolutionary and Ecologycal Dynamicsw - Schoener - Science-2011Документ5 страниц3-The Newest Synthesis - Understanding The Interplay of Evolutionary and Ecologycal Dynamicsw - Schoener - Science-2011Uriel RodríguezОценок пока нет
- Physiological Ecology of Tropical PlantsДокумент466 страницPhysiological Ecology of Tropical Plantsfernando1garc1a-1100% (2)
- Urata - Etal - Larval Development of The Oriental Lancelet - BranchiostomaДокумент11 страницUrata - Etal - Larval Development of The Oriental Lancelet - BranchiostomaRochyRiquelmeОценок пока нет
- Biology and Control of Invasive FishesДокумент278 страницBiology and Control of Invasive FishesHaliliОценок пока нет
- Molluscan Studies: Journal ofДокумент19 страницMolluscan Studies: Journal ofWidi SetyogatiОценок пока нет
- Deuterostomia Wanninger2015Документ220 страницDeuterostomia Wanninger2015Isabella Maria Trujillo PulgarОценок пока нет
- Anatomical Studies On The Cranial Nerves and Cranial Ganglia of Anguilla Anguilla (Linnaeus, 1758) Eye Muscle NervesДокумент21 страницаAnatomical Studies On The Cranial Nerves and Cranial Ganglia of Anguilla Anguilla (Linnaeus, 1758) Eye Muscle NervesIJAR JOURNALОценок пока нет
- 18 Sigwart Sumner Rooney Chapter 18Документ19 страниц18 Sigwart Sumner Rooney Chapter 18Haniza SaraОценок пока нет
- Digestive Function of Different BirdsДокумент27 страницDigestive Function of Different BirdsFiroz RezaОценок пока нет
- The Behavioural Ecology of The Komodo MonitorДокумент3 страницыThe Behavioural Ecology of The Komodo MonitorDed RanggaОценок пока нет
- Say Hello To Our Little Friends: ZOO br00011Документ3 страницыSay Hello To Our Little Friends: ZOO br00011Andjela RoganovicОценок пока нет
- PatelDivya - Week 2 AssignmentДокумент6 страницPatelDivya - Week 2 AssignmentDivya PatelОценок пока нет
- (PDF) PoriferaДокумент42 страницы(PDF) Poriferapm102382Оценок пока нет
- Finite Element Analysis of Ungulate Jaws Can Mode PDFДокумент16 страницFinite Element Analysis of Ungulate Jaws Can Mode PDFNonoОценок пока нет
- 2021 IAPO Bulletin 2021-2 5 ReshamДокумент5 страниц2021 IAPO Bulletin 2021-2 5 ReshamNećeš SadaОценок пока нет
- Walker - Argulus: Ecology of A Fish PestДокумент192 страницыWalker - Argulus: Ecology of A Fish Pestsadida anindyaОценок пока нет
- Abrusan - 2003 - Morphological Vaiation in LeptodoraДокумент6 страницAbrusan - 2003 - Morphological Vaiation in LeptodoraDavid CarrozzoОценок пока нет
- Anatomy of Teleost Fish Immune Structures and Organs: Håvard Bjørgen Erling Olaf KoppangДокумент11 страницAnatomy of Teleost Fish Immune Structures and Organs: Håvard Bjørgen Erling Olaf KoppangBayuОценок пока нет
- Diniz e Brito - o Carismático Tamanduá BandeiraДокумент8 страницDiniz e Brito - o Carismático Tamanduá BandeiraGabriel MarquesОценок пока нет
- Nunes de Almeida Et Al 2021Документ16 страницNunes de Almeida Et Al 2021carlitos salazarОценок пока нет
- Handbook of The Mammals of The WorldДокумент3 страницыHandbook of The Mammals of The WorldSunshine PowerОценок пока нет
- Laaka Et Al., 2003. Dispersal of Asexual Propagules in BryophytesДокумент13 страницLaaka Et Al., 2003. Dispersal of Asexual Propagules in BryophytesPepito GrilloОценок пока нет
- Tooth Wear As A Means To Quantify Intra-Specific Variations in Diet and Chewing Movements - Calandra Et Al 2016Документ10 страницTooth Wear As A Means To Quantify Intra-Specific Variations in Diet and Chewing Movements - Calandra Et Al 2016Bogdan HaiducОценок пока нет
- Color vision in marine mammals reviewДокумент16 страницColor vision in marine mammals reviewYuliana PurnamasariОценок пока нет
- The Evolution of Early Vertebrate Photoreceptors: ReviewДокумент16 страницThe Evolution of Early Vertebrate Photoreceptors: ReviewJ.D. NobleОценок пока нет
- 058 2016 Ferrando Et Al Polar BiologyДокумент13 страниц058 2016 Ferrando Et Al Polar BiologyharisankarhsОценок пока нет
- Ecology of BatsДокумент15 страницEcology of BatspomajoluОценок пока нет
- EndolimaxДокумент24 страницыEndolimaxconheoconОценок пока нет
- Quantifying Tidal Movements of The Shore Crab Carcinus Maenas On To Complex Epibenthic Bivalve HabitatsДокумент15 страницQuantifying Tidal Movements of The Shore Crab Carcinus Maenas On To Complex Epibenthic Bivalve HabitatsbambilovespascalОценок пока нет
- Further Structures in The Jaw Apparatus of Limnognathia Maerski (Micrognathozoa), With Notes On The Phylogeny of The GnathiferaДокумент15 страницFurther Structures in The Jaw Apparatus of Limnognathia Maerski (Micrognathozoa), With Notes On The Phylogeny of The GnathiferaEduardo ClaudesОценок пока нет
- Experiments: Reappraisal Gram-Negative Sepsis?Документ1 страницаExperiments: Reappraisal Gram-Negative Sepsis?Iuliana MogaОценок пока нет
- Paper GastropodaДокумент10 страницPaper GastropodaDrbernhardОценок пока нет
- Macffaen 2001. Primary and Secondary Endosymbiosis and The OriginДокумент10 страницMacffaen 2001. Primary and Secondary Endosymbiosis and The OriginGabyОценок пока нет
- Animal Structure and HabitesДокумент3 страницыAnimal Structure and HabitesVictor informaticoОценок пока нет
- V. J. Chapman M.a., Ph.D. (Cantab.), D. J. Chapman B.sc. (Auck.), Ph.D. (Cal.) (Auth.) - The Algae-Palgrave Macmillan UK (1973)Документ511 страницV. J. Chapman M.a., Ph.D. (Cantab.), D. J. Chapman B.sc. (Auck.), Ph.D. (Cal.) (Auth.) - The Algae-Palgrave Macmillan UK (1973)MichelОценок пока нет
- The Paranasal Sinuses and Other Enigmas - An Aquatic Evolutionary TheoryДокумент12 страницThe Paranasal Sinuses and Other Enigmas - An Aquatic Evolutionary TheoryBrookОценок пока нет
- Evolution of The Mammalian Neck From Developmental, Morpho-Functional, and Paleontological PerspectivesДокумент11 страницEvolution of The Mammalian Neck From Developmental, Morpho-Functional, and Paleontological PerspectivesKo KaОценок пока нет
- Ticks and Tick-Borne DiseasesДокумент8 страницTicks and Tick-Borne DiseasesLørïïs SïerraОценок пока нет
- Illustrated Summary of Chiton TermsДокумент25 страницIllustrated Summary of Chiton TermsDerrick Scott FullerОценок пока нет
- The Zebrafish Issue of DevelopmentДокумент5 страницThe Zebrafish Issue of DevelopmentKarthik ShettyОценок пока нет
- Handbook of The Mammals of The WorldДокумент3 страницыHandbook of The Mammals of The Worldadriani2003Оценок пока нет
- BRANCHFIELD 2016 - A Three-Dimensional Study of Alveologenesis in Mouse LungДокумент13 страницBRANCHFIELD 2016 - A Three-Dimensional Study of Alveologenesis in Mouse LungRafael JustinoОценок пока нет
- Bottlenose Dolphin Research PaperДокумент7 страницBottlenose Dolphin Research Papermadywedykul2100% (1)
- Despre PisiciДокумент25 страницDespre PisiciŞerban DobrincuОценок пока нет
- guide83Документ14 страницguide83DC NUPОценок пока нет
- Into The Brains of Whales: Mark Peter SimmondsДокумент14 страницInto The Brains of Whales: Mark Peter Simmondsapi-3806979Оценок пока нет
- Ear Structures of The Naked Mole-Rat, (Rodentia: Bathyergidae)Документ28 страницEar Structures of The Naked Mole-Rat, (Rodentia: Bathyergidae)Mellya RizkiОценок пока нет
- Plastic Response To Pond Drying in Tadpoles Rana Temporaria: Tests of Cost ModelsДокумент16 страницPlastic Response To Pond Drying in Tadpoles Rana Temporaria: Tests of Cost Modelstorrid64Оценок пока нет
- Cheetah 2014 Behavior EcoДокумент9 страницCheetah 2014 Behavior EcoHS RazОценок пока нет
- 2015 KlugHoffmann AmmonoidSepta 3Документ47 страниц2015 KlugHoffmann AmmonoidSepta 3Rafael Roldán de la RúaОценок пока нет
- Journal Homepage: - : IntroductionДокумент12 страницJournal Homepage: - : IntroductionIJAR JOURNALОценок пока нет
- Comments on the Taxonomic Status of Apodemus peninsulae, with Description of a New Subspecies from North ChinaОт EverandComments on the Taxonomic Status of Apodemus peninsulae, with Description of a New Subspecies from North ChinaОценок пока нет
- Advances in Genomics of Bony FishДокумент13 страницAdvances in Genomics of Bony FishBio HafidОценок пока нет
- European Neogene Mammal ChronologyДокумент9 страницEuropean Neogene Mammal ChronologyBogdan HaiducОценок пока нет
- Mammalian Teeth 03 Schulz KornasДокумент10 страницMammalian Teeth 03 Schulz KornasBogdan HaiducОценок пока нет
- Sheffield Edge Tool MakersДокумент48 страницSheffield Edge Tool MakersnikkicluОценок пока нет
- Cifelli Ed 1996Документ41 страницаCifelli Ed 1996Bogdan HaiducОценок пока нет
- Gr. Cobalcescu Annual Scientific Symposium - en 2019Документ2 страницыGr. Cobalcescu Annual Scientific Symposium - en 2019Bogdan HaiducОценок пока нет
- Tooth Wear As A Means To Quantify Intra-Specific Variations in Diet and Chewing Movements - Calandra Et Al 2016Документ10 страницTooth Wear As A Means To Quantify Intra-Specific Variations in Diet and Chewing Movements - Calandra Et Al 2016Bogdan HaiducОценок пока нет
- Haiduc Et Al. 2018Документ8 страницHaiduc Et Al. 2018Bogdan HaiducОценок пока нет
- PastmanualДокумент205 страницPastmanualSheetal BhosaleОценок пока нет
- The History of Mesowear - A Review, Ackermans 2020Документ17 страницThe History of Mesowear - A Review, Ackermans 2020Bogdan HaiducОценок пока нет
- Dental Microwear Texture Analysis: Technical ConsiderationsДокумент11 страницDental Microwear Texture Analysis: Technical ConsiderationsBogdan HaiducОценок пока нет
- Tooth Wear As A Means To Quantify Intra-Specific Variations in Diet and Chewing Movements - Calandra Et Al 2016Документ10 страницTooth Wear As A Means To Quantify Intra-Specific Variations in Diet and Chewing Movements - Calandra Et Al 2016Bogdan HaiducОценок пока нет
- Kelley & Gao (2012) PNAS - Yuanmou Cranium PDFДокумент4 страницыKelley & Gao (2012) PNAS - Yuanmou Cranium PDFBogdan HaiducОценок пока нет
- Kelley & Gao (2012) PNAS (SI) - Yuanmou CraniumДокумент3 страницыKelley & Gao (2012) PNAS (SI) - Yuanmou CraniumBogdan HaiducОценок пока нет
- Scott Et Al (2009) AJPA - Ouranopithecus Sexual DimorphismДокумент12 страницScott Et Al (2009) AJPA - Ouranopithecus Sexual DimorphismBogdan HaiducОценок пока нет
- Hershkovitz Et Al (1997) JHE - Sivapithecus Oral BacteriaДокумент6 страницHershkovitz Et Al (1997) JHE - Sivapithecus Oral BacteriaBogdan HaiducОценок пока нет
- Baigusheva Et Al - Tooth of Early Generations - QI - 2016Документ14 страницBaigusheva Et Al - Tooth of Early Generations - QI - 2016Bogdan HaiducОценок пока нет
- Pages From Ward Et Al (2000) Science - EquatoriusДокумент1 страницаPages From Ward Et Al (2000) Science - EquatoriusBogdan HaiducОценок пока нет
- Paleoenvironment in East Java During The Last 25,000 Years As Inferred From Bovid and Cervid Dental Wear Analyses - 2016 Rivals Et AlДокумент12 страницPaleoenvironment in East Java During The Last 25,000 Years As Inferred From Bovid and Cervid Dental Wear Analyses - 2016 Rivals Et AlBogdan HaiducОценок пока нет
- How To Install R PDFДокумент2 страницыHow To Install R PDFBogdan HaiducОценок пока нет
- Kelley & Alpagut (1999) JHE - Pasalar CaninesДокумент7 страницKelley & Alpagut (1999) JHE - Pasalar CaninesBogdan HaiducОценок пока нет
- Registration Form SPR 2017Документ1 страницаRegistration Form SPR 2017Bogdan HaiducОценок пока нет
- 2016 Impact Factor (JCR)Документ210 страниц2016 Impact Factor (JCR)dipsiiiОценок пока нет
- Why Does Micromorphology Have Such Little Traction in Geoarchaeology - Aldeias Et Al 2016Документ11 страницWhy Does Micromorphology Have Such Little Traction in Geoarchaeology - Aldeias Et Al 2016Bogdan HaiducОценок пока нет
- Phytoliths - A Comprehensive Guide For Archaeologists and PaleoecologistsДокумент2 страницыPhytoliths - A Comprehensive Guide For Archaeologists and PaleoecologistsBogdan HaiducОценок пока нет
- Kelley & Etler (1989) AJP - Lufeng Species NumberДокумент20 страницKelley & Etler (1989) AJP - Lufeng Species NumberBogdan HaiducОценок пока нет
- Evolution and Palaeobiology of PterosaursДокумент2 страницыEvolution and Palaeobiology of PterosaursBogdan HaiducОценок пока нет
- Comparison of Switching Strategies For An Ultrasonic Cleaner PDFДокумент5 страницComparison of Switching Strategies For An Ultrasonic Cleaner PDFBogdan HaiducОценок пока нет
- First Cenozoic BirdДокумент8 страницFirst Cenozoic BirdBogdan HaiducОценок пока нет
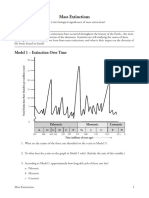
- Mass ExtinctionsДокумент6 страницMass ExtinctionsJacqueline50% (2)
- Strat Paper Complete PDFДокумент25 страницStrat Paper Complete PDFAmie WhitlockОценок пока нет
- Historical Geology (6th Ed) - R. Wicander & J. MonroeДокумент12 страницHistorical Geology (6th Ed) - R. Wicander & J. Monroemach20_aardvark80640% (1)
- Dating of FossilsДокумент23 страницыDating of FossilsshieldОценок пока нет
- Hominid/Human Evolution: Geology 331 PaleontologyДокумент109 страницHominid/Human Evolution: Geology 331 PaleontologyIsrael Gil Aguilar LacayОценок пока нет
- Biology NIOSДокумент927 страницBiology NIOSApshar RizwanОценок пока нет
- Clovis First TheoryДокумент3 страницыClovis First Theoryapi-325886206Оценок пока нет
- NouradiniДокумент56 страницNouradiniDavi AhmadОценок пока нет
- Jurassic Parts RulesДокумент16 страницJurassic Parts RulesSunnyОценок пока нет
- The Lost World of GiantsДокумент38 страницThe Lost World of Giantsmanticon01100% (2)
- Geology of PalawanДокумент6 страницGeology of PalawanDaniel Morabe100% (1)
- Out of Eden WalkДокумент2 страницыOut of Eden WalkGeorgian National MuseumОценок пока нет
- 185 Walter Et Al. 2005 - Journal of South American Earth SciencesДокумент11 страниц185 Walter Et Al. 2005 - Journal of South American Earth SciencesswithiodsОценок пока нет
- 3.trace FossilДокумент104 страницы3.trace FossilAldo IDОценок пока нет
- LADGASHTДокумент2 страницыLADGASHTEjaz ul Haq kakarОценок пока нет
- NO Class Type Description Location Lot Area Floor Area Selling Price Status Metro Manila CaloocanДокумент28 страницNO Class Type Description Location Lot Area Floor Area Selling Price Status Metro Manila Caloocanupg632Оценок пока нет
- Chapter 22 Descent With Modification A Darwinian View of LifeДокумент15 страницChapter 22 Descent With Modification A Darwinian View of Life蔡旻珊Оценок пока нет
- UKS2 DF BLKF Dinosaur Species S2 Resource1Документ5 страницUKS2 DF BLKF Dinosaur Species S2 Resource1Chris FalokunОценок пока нет
- MesosaurusДокумент9 страницMesosaurusApple PanganibanОценок пока нет
- Bangladesh StratigraphyДокумент19 страницBangladesh StratigraphyMahfuz Islam MeghОценок пока нет
- Fossil QuercusДокумент68 страницFossil QuercusAlexandru Sabin BadarauОценок пока нет
- Fishes - Golden Guide 1987 PDFДокумент164 страницыFishes - Golden Guide 1987 PDFConifor100% (2)
- Athreya and Ackermann 2018 Colonialism Narratives Human Origins Asia AfricaДокумент33 страницыAthreya and Ackermann 2018 Colonialism Narratives Human Origins Asia Africamoni_ayalauahОценок пока нет
- Stratigraphy Upper Indus BasinДокумент39 страницStratigraphy Upper Indus BasinAwais Khattak100% (1)
- Sistema Petrolero Del Campo GullfaksДокумент8 страницSistema Petrolero Del Campo GullfaksPepe MtzОценок пока нет
- What Is Plate TectonicsДокумент4 страницыWhat Is Plate TectonicsAkwa ManОценок пока нет
- Understanding Geologic Time 6 8 PDFДокумент5 страницUnderstanding Geologic Time 6 8 PDFAubrey CapolinasОценок пока нет
- Ug Anthropology Syllabus WbsuДокумент15 страницUg Anthropology Syllabus WbsuARPITA DUTTAОценок пока нет
- The Middle-Upper Paleolithic TransitionДокумент9 страницThe Middle-Upper Paleolithic TransitionKerim SuljevićОценок пока нет
- Ebridianos FósilesДокумент116 страницEbridianos FósilesFernando Ureña PlazaОценок пока нет