Вам также может понравиться
- Academic MisconductДокумент38 страницAcademic MisconductSisca Rizkia ArifiantiОценок пока нет
- 2014 Vol9 (12) No1 pg88-93Документ6 страниц2014 Vol9 (12) No1 pg88-93Sisca Rizkia ArifiantiОценок пока нет
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceОт EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceРейтинг: 4 из 5 звезд4/5 (894)
- Sik Gigi Mayang 23 April 2016Документ265 страницSik Gigi Mayang 23 April 2016Sisca Rizkia ArifiantiОценок пока нет
- The Yellow House: A Memoir (2019 National Book Award Winner)От EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Рейтинг: 4 из 5 звезд4/5 (98)
- Research Methods 3Документ35 страницResearch Methods 3Sisca Rizkia ArifiantiОценок пока нет
- The Little Book of Hygge: Danish Secrets to Happy LivingОт EverandThe Little Book of Hygge: Danish Secrets to Happy LivingРейтинг: 3.5 из 5 звезд3.5/5 (399)
- Cabt 13 I 2 P 258Документ5 страницCabt 13 I 2 P 258Sisca Rizkia ArifiantiОценок пока нет
- Coinfection With Herpes Simplex Virus Type 2Документ8 страницCoinfection With Herpes Simplex Virus Type 2Sisca Rizkia ArifiantiОценок пока нет
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeОт EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeРейтинг: 4 из 5 звезд4/5 (5794)
- Long-Term Valacyclovir Suppressive Treatment After Herpes Simplex Virus Type 2 MeningitisДокумент10 страницLong-Term Valacyclovir Suppressive Treatment After Herpes Simplex Virus Type 2 MeningitisSisca Rizkia ArifiantiОценок пока нет
- Never Split the Difference: Negotiating As If Your Life Depended On ItОт EverandNever Split the Difference: Negotiating As If Your Life Depended On ItРейтинг: 4.5 из 5 звезд4.5/5 (838)
- Prevalent Herpes Simplex Virus Type 2 Infection Is AssociatedДокумент6 страницPrevalent Herpes Simplex Virus Type 2 Infection Is AssociatedSisca Rizkia ArifiantiОценок пока нет
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureОт EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureРейтинг: 4.5 из 5 звезд4.5/5 (474)
- Nej Mo A 1410649Документ10 страницNej Mo A 1410649Sisca Rizkia ArifiantiОценок пока нет
- Ijm 4 204Документ6 страницIjm 4 204Sisca Rizkia ArifiantiОценок пока нет
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryОт EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryРейтинг: 3.5 из 5 звезд3.5/5 (231)
- Herpes Simplex Virus Suppression WithДокумент9 страницHerpes Simplex Virus Suppression WithSisca Rizkia ArifiantiОценок пока нет
- C H C /C (C L) : Hronic Yperplastic Andidosis Andidiasis Andidal EukoplakiaДокумент15 страницC H C /C (C L) : Hronic Yperplastic Andidosis Andidiasis Andidal EukoplakiaSisca Rizkia ArifiantiОценок пока нет
- The Emperor of All Maladies: A Biography of CancerОт EverandThe Emperor of All Maladies: A Biography of CancerРейтинг: 4.5 из 5 звезд4.5/5 (271)
- Review Artikel Kelompok 2Документ10 страницReview Artikel Kelompok 2Yuanita Hasna RahmadhaniОценок пока нет
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreОт EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreРейтинг: 4 из 5 звезд4/5 (1090)
- Geographic Tongue and Fissured Tongue in 348 Patients With Psoriasis: Correlation With Disease SeverityДокумент8 страницGeographic Tongue and Fissured Tongue in 348 Patients With Psoriasis: Correlation With Disease SeveritypancholarpancholarОценок пока нет
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyОт EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyРейтинг: 3.5 из 5 звезд3.5/5 (2219)
- 2686 PDFДокумент10 страниц2686 PDFSisca Rizkia ArifiantiОценок пока нет
- Team of Rivals: The Political Genius of Abraham LincolnОт EverandTeam of Rivals: The Political Genius of Abraham LincolnРейтинг: 4.5 из 5 звезд4.5/5 (234)
- 2014 Vol9 (12) No1 pg88-93Документ6 страниц2014 Vol9 (12) No1 pg88-93Sisca Rizkia ArifiantiОценок пока нет
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersОт EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersРейтинг: 4.5 из 5 звезд4.5/5 (344)
- C H C /C (C L) : Hronic Yperplastic Andidosis Andidiasis Andidal EukoplakiaДокумент15 страницC H C /C (C L) : Hronic Yperplastic Andidosis Andidiasis Andidal EukoplakiaSisca Rizkia ArifiantiОценок пока нет
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaОт EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaРейтинг: 4.5 из 5 звезд4.5/5 (265)
- Immune Response and Candidal Colonisation in Denture Associated Stomatitis 2155 9899.1000178Документ7 страницImmune Response and Candidal Colonisation in Denture Associated Stomatitis 2155 9899.1000178Sisca Rizkia ArifiantiОценок пока нет
- Chronic Periadontitis As PDFДокумент6 страницChronic Periadontitis As PDFAhtarunnisa Fauzia HanifaОценок пока нет
- Ijm 4 204Документ6 страницIjm 4 204Sisca Rizkia ArifiantiОценок пока нет
- 2686 PDFДокумент10 страниц2686 PDFSisca Rizkia ArifiantiОценок пока нет
- B.ing GJДокумент13 страницB.ing GJSisca Rizkia ArifiantiОценок пока нет
- Case Report: A.N Atypical Pathway of Infection in An Adolescent W TH A Deep Neck Space AbscessДокумент4 страницыCase Report: A.N Atypical Pathway of Infection in An Adolescent W TH A Deep Neck Space AbscessSisca Rizkia ArifiantiОценок пока нет
- The Unwinding: An Inner History of the New AmericaОт EverandThe Unwinding: An Inner History of the New AmericaРейтинг: 4 из 5 звезд4/5 (45)
- Prima HidayaningtyasДокумент13 страницPrima Hidayaningtyasv3_nanaОценок пока нет
- Print St2 AnatomiДокумент1 страницаPrint St2 AnatomiSisca Rizkia ArifiantiОценок пока нет
- StdsДокумент14 страницStdsJumar Divinagracia DimpasОценок пока нет
- Sexually Transmitted Disease and HomeopathyДокумент5 страницSexually Transmitted Disease and HomeopathyMihaela Ghica100% (1)
- Viral ConjunctivitisДокумент30 страницViral ConjunctivitisJoane Chan LimОценок пока нет
- Atlas and Synopsis of Clinical DermatologyДокумент1 146 страницAtlas and Synopsis of Clinical DermatologyAnda Madalina Zaharia100% (6)
- Phytochemical Analysis and in Vitro Evaluation of The Biological Activity Against Herpes Simplex Virus Type 1 (HSV-1) of Cedrus Libani A. RichДокумент5 страницPhytochemical Analysis and in Vitro Evaluation of The Biological Activity Against Herpes Simplex Virus Type 1 (HSV-1) of Cedrus Libani A. RichAdibah BahtiarОценок пока нет
- Viral Infections in Sudden Hearing Loss. Do We Have Enough Evidence?Документ4 страницыViral Infections in Sudden Hearing Loss. Do We Have Enough Evidence?VersoОценок пока нет
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)От EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Рейтинг: 4.5 из 5 звезд4.5/5 (119)
- The$Guide$for$Newly$ Graduated$doctor$ $ $ $ $ Collected$By:$ DR - soran$Mohammed$$Gharib$ M.B.CH.B$ $ $ $$ $ $Документ210 страницThe$Guide$for$Newly$ Graduated$doctor$ $ $ $ $ Collected$By:$ DR - soran$Mohammed$$Gharib$ M.B.CH.B$ $ $ $$ $ $Raghib KurdiОценок пока нет
- Chapter 28 - InfectionsДокумент6 страницChapter 28 - InfectionsEunice CortésОценок пока нет
- Nematodes & Infections of The SkinДокумент39 страницNematodes & Infections of The SkinHannah LaputОценок пока нет
- Advances in Neonatal InfectionsДокумент5 страницAdvances in Neonatal InfectionsElizabeth HendersonОценок пока нет
- Appendicitis Menigitis Shingles (Mayo Clinic Def)Документ11 страницAppendicitis Menigitis Shingles (Mayo Clinic Def)Lucila LugoОценок пока нет
- Bacterial Infections of the Mouth and BeyondДокумент1 страницаBacterial Infections of the Mouth and BeyondTaif SalimОценок пока нет
- Skin and Eye Infection MicpДокумент17 страницSkin and Eye Infection MicpMarc Kevin MendozaОценок пока нет
- Common Viral Infections Including Corona (Covid Infection) in Children - A ReviewДокумент10 страницCommon Viral Infections Including Corona (Covid Infection) in Children - A ReviewIJAR JOURNALОценок пока нет
- Premarital SexДокумент6 страницPremarital SexArcon AlvarОценок пока нет
- LYSOVITДокумент2 страницыLYSOVITomairОценок пока нет
- Lemone/Burke/Bauldoff/Gubrud, Medical-Surgical Nursing 5Th Edition Test BankДокумент41 страницаLemone/Burke/Bauldoff/Gubrud, Medical-Surgical Nursing 5Th Edition Test Banknurse homeОценок пока нет
- Knust Lect Infectious Disease IДокумент8 страницKnust Lect Infectious Disease IAnastasiafynnОценок пока нет
- Herpes ZosterДокумент12 страницHerpes ZosterCharlz ZipaganОценок пока нет
- Viral PathogenesisДокумент14 страницViral PathogenesisMohammed Yousif mzoriОценок пока нет
- Cytodiagnosis in DermatologyДокумент5 страницCytodiagnosis in DermatologyDipti MathiasОценок пока нет
- Cytotoxic Killing Cell Apoptosis. F Liver Inflammation Liver Damage 1-6 MONTHSДокумент21 страницаCytotoxic Killing Cell Apoptosis. F Liver Inflammation Liver Damage 1-6 MONTHSKyla MendozaОценок пока нет
- Booklet 2018Документ34 страницыBooklet 2018BaluErnestoОценок пока нет
- Tea Tree OilДокумент66 страницTea Tree OilGiselda FarmasiОценок пока нет
- STD - Infection That Are Passed From One Person To Another Through Sexual Contact. The Causes of StdsДокумент3 страницыSTD - Infection That Are Passed From One Person To Another Through Sexual Contact. The Causes of StdsAngela BeatriceОценок пока нет
- Infectous DiseaseДокумент9 страницInfectous DiseasejamikawasОценок пока нет
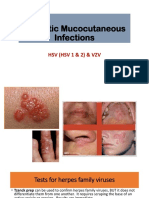
- Herpetic Mucocutaneous Infections: HSV (HSV 1 & 2) & VZVДокумент32 страницыHerpetic Mucocutaneous Infections: HSV (HSV 1 & 2) & VZVJaneОценок пока нет
- Olive Leaf ExtractДокумент9 страницOlive Leaf ExtractCristina100% (1)
- Herpes Genitalis: Causes, Symptoms, TreatmentДокумент13 страницHerpes Genitalis: Causes, Symptoms, TreatmentJimson Marc DuranОценок пока нет
- Report HPV, Shingles and Chicken PoxДокумент4 страницыReport HPV, Shingles and Chicken PoxJarold PerezОценок пока нет
- LIT: Life Ignition Tools: Use Nature's Playbook to Energize Your Brain, Spark Ideas, and Ignite ActionОт EverandLIT: Life Ignition Tools: Use Nature's Playbook to Energize Your Brain, Spark Ideas, and Ignite ActionРейтинг: 4 из 5 звезд4/5 (402)
- Mindset by Carol S. Dweck - Book Summary: The New Psychology of SuccessОт EverandMindset by Carol S. Dweck - Book Summary: The New Psychology of SuccessРейтинг: 4.5 из 5 звезд4.5/5 (327)
- The Age of Magical Overthinking: Notes on Modern IrrationalityОт EverandThe Age of Magical Overthinking: Notes on Modern IrrationalityРейтинг: 4 из 5 звезд4/5 (13)
- The Ultimate Guide To Memory Improvement TechniquesОт EverandThe Ultimate Guide To Memory Improvement TechniquesРейтинг: 5 из 5 звезд5/5 (34)
- Techniques Exercises And Tricks For Memory ImprovementОт EverandTechniques Exercises And Tricks For Memory ImprovementРейтинг: 4.5 из 5 звезд4.5/5 (40)
- The Ritual Effect: From Habit to Ritual, Harness the Surprising Power of Everyday ActionsОт EverandThe Ritual Effect: From Habit to Ritual, Harness the Surprising Power of Everyday ActionsРейтинг: 3.5 из 5 звезд3.5/5 (3)