Вам также может понравиться
- The Yellow House: A Memoir (2019 National Book Award Winner)От EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Рейтинг: 4 из 5 звезд4/5 (98)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceОт EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceРейтинг: 4 из 5 звезд4/5 (895)
- P 2doДокумент2 страницыP 2doCarlos MontúfarОценок пока нет
- P 2doДокумент2 страницыP 2doCarlos MontúfarОценок пока нет
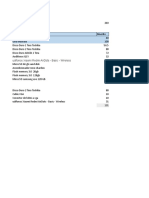
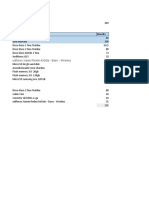
- PresupuestoДокумент3 страницыPresupuestoCarlos MontúfarОценок пока нет
- P 3roДокумент2 страницыP 3roCarlos MontúfarОценок пока нет
- PresupuestoДокумент3 страницыPresupuestoCarlos MontúfarОценок пока нет
- P 3roДокумент2 страницыP 3roCarlos MontúfarОценок пока нет
- 12Документ3 страницы12Carlos MontúfarОценок пока нет
- Artículo 2Документ2 страницыArtículo 2yessОценок пока нет
- The Navier-Stokes Equations: 14.1 Conservation LawsДокумент14 страницThe Navier-Stokes Equations: 14.1 Conservation LawsCarlos MontúfarОценок пока нет
- ProgramaДокумент5 страницProgramaCarlos MontúfarОценок пока нет
- Re Re: 10 TurbulentoДокумент2 страницыRe Re: 10 TurbulentoCarlos MontúfarОценок пока нет
- Moody Chart SolverДокумент2 страницыMoody Chart SolverHernâniCruzОценок пока нет
- Gas TuberiasДокумент1 страницаGas TuberiasCarlos MontúfarОценок пока нет
- OperativaДокумент18 страницOperativaCarlos MontúfarОценок пока нет
- MichaelДокумент29 страницMichaelCarlos MontúfarОценок пока нет
- MM Kinetics No.4 PDFДокумент6 страницMM Kinetics No.4 PDFCarlos MontúfarОценок пока нет
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeОт EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeРейтинг: 4 из 5 звезд4/5 (5794)
- The Little Book of Hygge: Danish Secrets to Happy LivingОт EverandThe Little Book of Hygge: Danish Secrets to Happy LivingРейтинг: 3.5 из 5 звезд3.5/5 (399)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaОт EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaРейтинг: 4.5 из 5 звезд4.5/5 (266)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureОт EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureРейтинг: 4.5 из 5 звезд4.5/5 (474)
- Never Split the Difference: Negotiating As If Your Life Depended On ItОт EverandNever Split the Difference: Negotiating As If Your Life Depended On ItРейтинг: 4.5 из 5 звезд4.5/5 (838)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryОт EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryРейтинг: 3.5 из 5 звезд3.5/5 (231)
- The Emperor of All Maladies: A Biography of CancerОт EverandThe Emperor of All Maladies: A Biography of CancerРейтинг: 4.5 из 5 звезд4.5/5 (271)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyОт EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyРейтинг: 3.5 из 5 звезд3.5/5 (2259)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersОт EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersРейтинг: 4.5 из 5 звезд4.5/5 (344)
- Team of Rivals: The Political Genius of Abraham LincolnОт EverandTeam of Rivals: The Political Genius of Abraham LincolnРейтинг: 4.5 из 5 звезд4.5/5 (234)
- The Unwinding: An Inner History of the New AmericaОт EverandThe Unwinding: An Inner History of the New AmericaРейтинг: 4 из 5 звезд4/5 (45)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreОт EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreРейтинг: 4 из 5 звезд4/5 (1090)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)От EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Рейтинг: 4.5 из 5 звезд4.5/5 (121)
- Net Force and Acceleration Practice Worksheet KEY 4Документ1 страницаNet Force and Acceleration Practice Worksheet KEY 4brooo imlikeoveritОценок пока нет
- Tesis Damian TДокумент359 страницTesis Damian TpalfrancaОценок пока нет
- Final Exam Class 11 Maths Exam2020-2021 Rashmi PДокумент5 страницFinal Exam Class 11 Maths Exam2020-2021 Rashmi Pnabhijain9Оценок пока нет
- Excel - Compare Two ColumnsДокумент4 страницыExcel - Compare Two ColumnsSusmitaMandalОценок пока нет
- Temperaments and Tunings - A Guide For Lute PlayersДокумент7 страницTemperaments and Tunings - A Guide For Lute PlayerschkornarosОценок пока нет
- Alphabet TheoryДокумент9 страницAlphabet TheoryMahendra JayanОценок пока нет
- UTS Bahasa Inggris AnalisisДокумент4 страницыUTS Bahasa Inggris AnalisisCuapCuapIndyОценок пока нет
- LR (0) Parse AlgorithmДокумент12 страницLR (0) Parse AlgorithmTanmoy MondalОценок пока нет
- PHD Thesis JD - CR - GS - FinalДокумент185 страницPHD Thesis JD - CR - GS - FinalmvillabrОценок пока нет
- PDSP ArchitectureДокумент95 страницPDSP ArchitecturekrajasekarantutiОценок пока нет
- DAA Lab ProgramsДокумент26 страницDAA Lab ProgramsV ManjunathОценок пока нет
- Math9 Module 9Документ21 страницаMath9 Module 9Diane Reign MianaguaОценок пока нет
- Polymer Supply Chain Management: An Industry White PaperДокумент15 страницPolymer Supply Chain Management: An Industry White PaperAnkit AgrawalОценок пока нет
- Year 7 Angles and Shapes WorkbookДокумент37 страницYear 7 Angles and Shapes Workbookapi-291565828Оценок пока нет
- Research Proposal 1 PDFДокумент5 страницResearch Proposal 1 PDFMunem BushraОценок пока нет
- Yoshibash 1997 - Numerical Analysis of Edge SingularitiesДокумент22 страницыYoshibash 1997 - Numerical Analysis of Edge SingularitiesbrockmraОценок пока нет
- VNR Sir Profile PDFДокумент16 страницVNR Sir Profile PDFNaagDusaОценок пока нет
- Notes - Class X, Physics, - KinematicsДокумент9 страницNotes - Class X, Physics, - Kinematicscmaheswara-1Оценок пока нет
- Physics Paper 3 For SPM 2019 2020 - AnswerДокумент35 страницPhysics Paper 3 For SPM 2019 2020 - AnswerAzman SelamatОценок пока нет
- DSP-Chapter3 Student 08022017Документ49 страницDSP-Chapter3 Student 08022017Thành Vinh PhạmОценок пока нет
- Electrical Formulas ExplanationsДокумент16 страницElectrical Formulas ExplanationsJShearer100% (24)
- The Basics of Capital Budgeting Evaluating Cash FlowsДокумент59 страницThe Basics of Capital Budgeting Evaluating Cash FlowsBabasab Patil (Karrisatte)Оценок пока нет
- Matrix Mathematics Chemical Problems: ApplicationsДокумент9 страницMatrix Mathematics Chemical Problems: ApplicationsCarlosОценок пока нет
- ENGLISH - Subject Verb Agreement Grammar Revision Guide and Quick Quiz Powerpoint AustralianДокумент11 страницENGLISH - Subject Verb Agreement Grammar Revision Guide and Quick Quiz Powerpoint AustralianRogelio LadieroОценок пока нет
- Strategic Intervention Material - Speed and VelocityДокумент24 страницыStrategic Intervention Material - Speed and VelocityRhonnel Manatad Alburo98% (194)
- Real GasДокумент8 страницReal GasAmartya AnshumanОценок пока нет
- Concrete Technology (CE205) Concrete Technology Laboratory Report Academic Year (2019-2020)Документ10 страницConcrete Technology (CE205) Concrete Technology Laboratory Report Academic Year (2019-2020)Hawar AhmadОценок пока нет
- Area Extension QuestionsДокумент10 страницArea Extension QuestionsBob CrossОценок пока нет
- T Distribution and Test of Hypothesis Part I STEMДокумент30 страницT Distribution and Test of Hypothesis Part I STEMnicole quilangОценок пока нет