Вам также может понравиться
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeОт EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeРейтинг: 4 из 5 звезд4/5 (5794)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceОт EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceРейтинг: 4 из 5 звезд4/5 (895)
- The Yellow House: A Memoir (2019 National Book Award Winner)От EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Рейтинг: 4 из 5 звезд4/5 (98)
- The Little Book of Hygge: Danish Secrets to Happy LivingОт EverandThe Little Book of Hygge: Danish Secrets to Happy LivingРейтинг: 3.5 из 5 звезд3.5/5 (400)
- The Emperor of All Maladies: A Biography of CancerОт EverandThe Emperor of All Maladies: A Biography of CancerРейтинг: 4.5 из 5 звезд4.5/5 (271)
- Never Split the Difference: Negotiating As If Your Life Depended On ItОт EverandNever Split the Difference: Negotiating As If Your Life Depended On ItРейтинг: 4.5 из 5 звезд4.5/5 (838)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyОт EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyРейтинг: 3.5 из 5 звезд3.5/5 (2259)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureОт EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureРейтинг: 4.5 из 5 звезд4.5/5 (474)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryОт EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryРейтинг: 3.5 из 5 звезд3.5/5 (231)
- Team of Rivals: The Political Genius of Abraham LincolnОт EverandTeam of Rivals: The Political Genius of Abraham LincolnРейтинг: 4.5 из 5 звезд4.5/5 (234)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaОт EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaРейтинг: 4.5 из 5 звезд4.5/5 (266)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersОт EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersРейтинг: 4.5 из 5 звезд4.5/5 (345)
- The Unwinding: An Inner History of the New AmericaОт EverandThe Unwinding: An Inner History of the New AmericaРейтинг: 4 из 5 звезд4/5 (45)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreОт EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreРейтинг: 4 из 5 звезд4/5 (1090)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)От EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Рейтинг: 4.5 из 5 звезд4.5/5 (121)
- Anatomy and Physiology CH 4 To 7 Flash CardsДокумент19 страницAnatomy and Physiology CH 4 To 7 Flash Cardsmalenya1Оценок пока нет
- GynoeciumДокумент7 страницGynoeciumpdekraaijОценок пока нет
- EntomologyДокумент50 страницEntomologyapi-19960301100% (1)
- 201 - The Saggy Baggy Elephant - See, Hear, ReadДокумент28 страниц201 - The Saggy Baggy Elephant - See, Hear, ReadAndrew RukinОценок пока нет
- Extinct Birds 2nd EditionДокумент609 страницExtinct Birds 2nd EditionRanIruka100% (3)
- Gas Exchange in Animals: We Will Be Studying The Diversity of Adaptations For This Process in Four Animal GroupsДокумент42 страницыGas Exchange in Animals: We Will Be Studying The Diversity of Adaptations For This Process in Four Animal GroupsMiss IqbalОценок пока нет
- Palaeoniscoid Remains From The Lower Permian Pedra de Fogo Formation Parna Ba Basin Insights From General Morphology and HistologyДокумент12 страницPalaeoniscoid Remains From The Lower Permian Pedra de Fogo Formation Parna Ba Basin Insights From General Morphology and HistologyVictor FontesОценок пока нет
- Group The Animals According To Their Breathing StructuresДокумент14 страницGroup The Animals According To Their Breathing StructuresIlah GhazaliОценок пока нет
- Mock Ospe Me211Документ15 страницMock Ospe Me211sahirbuleОценок пока нет
- Apg IiiДокумент17 страницApg IiilokoОценок пока нет
- Flyer Manuale - CON WATERMARK PAGINE SINGOLEДокумент6 страницFlyer Manuale - CON WATERMARK PAGINE SINGOLEjscmtОценок пока нет
- Phylum Platyhelminthes Phylum Nematoda Phylum MolluscaДокумент44 страницыPhylum Platyhelminthes Phylum Nematoda Phylum MolluscaAngel FebiolaОценок пока нет
- Full Basic Psychopharmacology For Counselors and Psychotherapists 2Nd Edition Sinacola Test Bank PDFДокумент25 страницFull Basic Psychopharmacology For Counselors and Psychotherapists 2Nd Edition Sinacola Test Bank PDFwilliam.ely879100% (16)
- Larval Forms in EchinodermataДокумент6 страницLarval Forms in EchinodermatanikhilОценок пока нет
- Coastal Crabs A Guide To The Crabs of New Zealand 2015Документ55 страницCoastal Crabs A Guide To The Crabs of New Zealand 2015Carlos GarciaОценок пока нет
- 2 Mora y Robinson Loxocemus - LepidochelysДокумент2 страницы2 Mora y Robinson Loxocemus - LepidochelysJosé MoraОценок пока нет
- 001-111 mmv76 Gomon Choerodon 7 WebДокумент111 страниц001-111 mmv76 Gomon Choerodon 7 WebM DayatОценок пока нет
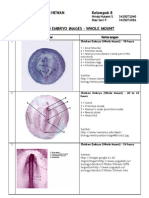
- Chick Embryo WMДокумент6 страницChick Embryo WMdhyan_ajjah67% (3)
- A New Teiid Lizard From The Late Cretaceous of The Haţeg Basin, Romania and Its Phylogenetic and Palaeobiogeographical RelationshipsДокумент20 страницA New Teiid Lizard From The Late Cretaceous of The Haţeg Basin, Romania and Its Phylogenetic and Palaeobiogeographical RelationshipsJ.D. NobleОценок пока нет
- Zoology McqsДокумент6 страницZoology McqsPrime RoseОценок пока нет
- Amazonas Nov Dic18Документ100 страницAmazonas Nov Dic18Sebastian GaunaОценок пока нет
- GeneticsДокумент13 страницGeneticsNina SapphireОценок пока нет
- For Connect With Final Answers Gen. Int 2 in Class Lecture 6 Gene Interaction - PART 2Документ33 страницыFor Connect With Final Answers Gen. Int 2 in Class Lecture 6 Gene Interaction - PART 2Anonymous Ef82K4xyОценок пока нет
- Cobb500 SF Breeder Management Chart Pounds PDFДокумент2 страницыCobb500 SF Breeder Management Chart Pounds PDFjai10141Оценок пока нет
- Identifying Birds by Their Beaks Lesson Plan PDFДокумент3 страницыIdentifying Birds by Their Beaks Lesson Plan PDFASHISH PADHY100% (1)
- Anatomy and Physiology 6th Edition Marieb Solutions ManualДокумент20 страницAnatomy and Physiology 6th Edition Marieb Solutions Manualstephenthanh1huo100% (28)
- Mammals in Sri LankaДокумент2 страницыMammals in Sri LankaVangeesa SumanasekaraОценок пока нет
- Dictionary of Plant Biology En-SpДокумент654 страницыDictionary of Plant Biology En-SpMelo2008100% (1)
- Class 11 Unit 1 Diversity in Living Organism Ch-1,2 (Plant and Animal Kingdom) NotesДокумент18 страницClass 11 Unit 1 Diversity in Living Organism Ch-1,2 (Plant and Animal Kingdom) NotesDhana Varshini maths biologyОценок пока нет
- List of AnimalДокумент11 страницList of AnimalTrần HảiОценок пока нет