Вам также может понравиться
- Qué Es MeiosisДокумент7 страницQué Es MeiosisYosguel DavidОценок пока нет
- Embriologia Falta CaratulaДокумент40 страницEmbriologia Falta CaratulaSaira Diaz AldanaОценок пока нет
- GametogénesisДокумент27 страницGametogénesisSergio Alejandro IbarraОценок пока нет
- GametosДокумент4 страницыGametosGrisel RodriguezОценок пока нет
- 2.2) Masculino PDFДокумент6 страниц2.2) Masculino PDFGiaan OrtizОценок пока нет
- El Sistema Genital MasculinoДокумент11 страницEl Sistema Genital MasculinoKarla Priscila AndradeОценок пока нет
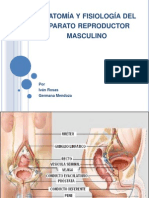
- Anatomía y Fisiología Del Aparato Reproductor MasculinoДокумент63 страницыAnatomía y Fisiología Del Aparato Reproductor MasculinoIvan RosasОценок пока нет
- Aparato Reproductor Del MachoДокумент10 страницAparato Reproductor Del MachoVanne Ag100% (1)
- EMBRIOLOGIAДокумент9 страницEMBRIOLOGIAAlejandra MartinezОценок пока нет
- TesticuloДокумент35 страницTesticuloLaura Angelica de PerezОценок пока нет
- Meiosis CurtisДокумент6 страницMeiosis CurtisMetzimx GuzmánОценок пока нет
- Mitosis, Meiosis y GametogenesisДокумент10 страницMitosis, Meiosis y GametogenesisBeatrizLinaresОценок пока нет
- Aparato Reproductor MasculinoДокумент11 страницAparato Reproductor MasculinoNair MartínezОценок пока нет
- E SpermatogenesisДокумент4 страницыE Spermatogenesispriscila castilloОценок пока нет
- Resumen Ross-Pawlina - Histología - Capitulo 22 - Sistema Genital MasculinoДокумент13 страницResumen Ross-Pawlina - Histología - Capitulo 22 - Sistema Genital Masculinovictoriaetchegoyen123Оценок пока нет
- Espermatogenesis - TrabajoДокумент12 страницEspermatogenesis - TrabajoJanella Tineo MedinaОценок пока нет
- Cromosomas Del CaninoДокумент8 страницCromosomas Del CaninoJose Adrian RodriguezОценок пока нет
- Meiosis y GamatogenesisДокумент5 страницMeiosis y GamatogenesisMaria Angeles QuesadaОценок пока нет
- GAMETOGÉNESIS - LangmanДокумент5 страницGAMETOGÉNESIS - LangmanViviana PalaciosОценок пока нет
- Embriologia 2Документ69 страницEmbriologia 2TamaraОценок пока нет
- Práctica 1 Morfo 2021Документ8 страницPráctica 1 Morfo 2021lyaОценок пока нет
- Marco Teórico: Generalidades Del Aparato Reproductor MasculinoДокумент18 страницMarco Teórico: Generalidades Del Aparato Reproductor MasculinoAndrea IriasОценок пока нет
- Atencion en Salud Materna Aparato ReproductorДокумент20 страницAtencion en Salud Materna Aparato Reproductorhenry ilaquita ccopaОценок пока нет
- Sistema Reproductor MasculinoДокумент135 страницSistema Reproductor MasculinoPaola Azucena SalasОценок пока нет
- Desarrollo EmbrionarioДокумент7 страницDesarrollo EmbrionarioKARLA ANDREA FLORES TORRESОценок пока нет
- Copia de Ciclo CelularДокумент4 страницыCopia de Ciclo CelularBianca CossioОценок пока нет
- GL OsarioДокумент4 страницыGL Osariojesus perezОценок пока нет
- Aparato Reproductor MasculinoДокумент8 страницAparato Reproductor Masculinoabril crespoОценок пока нет
- Resumen Final Embriología.Документ17 страницResumen Final Embriología.Nazareth PacayutОценок пока нет
- ESPERMATOGENESISДокумент15 страницESPERMATOGENESISMiriam E. MairimОценок пока нет
- MeiosisДокумент30 страницMeiosisSandra MilenaОценок пока нет
- MEIOSISДокумент30 страницMEIOSISJulio Eduardo CiodaroОценок пока нет
- Espermatogenesis Practica EmbriologiaДокумент9 страницEspermatogenesis Practica EmbriologiaJasminii Flor Llatas MontenegroОценок пока нет
- Division CelularДокумент51 страницаDivision CelularJosselin Jazmin Zanga QuispeОценок пока нет
- Embrio Odonto V2Документ22 страницыEmbrio Odonto V2cosadedubsОценок пока нет
- EspermatogénesisДокумент25 страницEspermatogénesisAdrianFonsecaMDОценок пока нет
- Gametogénesis I y IIДокумент19 страницGametogénesis I y IINotnow MaybelaterОценок пока нет
- EspermatogénesisДокумент5 страницEspermatogénesisXavier GuillenОценок пока нет
- Las Celulas SexualesДокумент52 страницыLas Celulas SexualesCindy MapulaОценок пока нет
- Histología GuíaДокумент22 страницыHistología GuíaAdi AguirreОценок пока нет
- Clase Histologia MasculinaДокумент59 страницClase Histologia MasculinaJorge Enrique Vera SagásteguiОценок пока нет
- Aparato Reproductor MasculinoДокумент5 страницAparato Reproductor MasculinoIzbeth TorresОценок пока нет
- Analisis Del Sistema Reproductor MasculinoДокумент2 страницыAnalisis Del Sistema Reproductor MasculinoAnderson BacajolОценок пока нет
- Gametognesis BiologiaДокумент15 страницGametognesis BiologiaJosue RebolledoОценок пока нет
- ESPERMATOGÉNESISДокумент4 страницыESPERMATOGÉNESISRosa Iriondo MendezОценок пока нет
- ReproducciónДокумент8 страницReproducciónSofia BoyeroОценок пока нет
- Aparatos ReproductoresДокумент38 страницAparatos ReproductoresAlisson CahuanaОценок пока нет
- Guias Octavo Cuarto PeriodoДокумент12 страницGuias Octavo Cuarto PeriodoJuan Jose Alvarez SierraОценок пока нет
- Sistema ResumidoДокумент24 страницыSistema ResumidoJorge RamirezОценок пока нет
- Histologia Del Aparato Reproductor MasculinoДокумент5 страницHistologia Del Aparato Reproductor MasculinoMauricio Leon100% (1)
- Aparato Reproductor Masculino33Документ4 страницыAparato Reproductor Masculino33Alexis Jhoan Oliveros DiazОценок пока нет
- Herencia Con Mitosis y MeiosisДокумент5 страницHerencia Con Mitosis y MeiosisMishell Hernández HernandezОценок пока нет
- Que Son Los CromosomasДокумент10 страницQue Son Los CromosomasIsidora Martínez GajardoОценок пока нет
- MeiosisДокумент4 страницыMeiosisYeimy Yulieth Ramirez MendezОценок пока нет
- SEM 7b MeiosisДокумент21 страницаSEM 7b MeiosisPatrick SayagoОценок пока нет
- Taller MeiosisДокумент6 страницTaller MeiosisTatiana LMОценок пока нет
- Gametogenesis Espermatogenesisi Ovogenesis InformeДокумент15 страницGametogenesis Espermatogenesisi Ovogenesis InformeBAUTISTA TERRONES GILMERОценок пока нет
- Anemia Para Todos: Medicina Para TodosОт EverandAnemia Para Todos: Medicina Para TodosРейтинг: 3 из 5 звезд3/5 (1)
- Funciones SuperioresДокумент8 страницFunciones Superiores1718h9798sОценок пока нет
- CONCIENCIAДокумент48 страницCONCIENCIA1718h9798sОценок пока нет
- Fármacos Utilizados en La Insuficiencia CardíacaДокумент13 страницFármacos Utilizados en La Insuficiencia Cardíaca1718h9798sОценок пока нет
- Expo FarmaДокумент13 страницExpo Farma1718h9798sОценок пока нет
- IntradermoreacciónДокумент2 страницыIntradermoreacción1718h9798sОценок пока нет
- Otros Exámenes ComplementariosДокумент7 страницOtros Exámenes Complementarios1718h9798sОценок пока нет
- PANCREATITISДокумент27 страницPANCREATITIS1718h9798sОценок пока нет
- Vía Extrapiramidal-Movimientos InvoluntariosДокумент3 страницыVía Extrapiramidal-Movimientos Involuntarios1718h9798sОценок пока нет
- Biopsia de PielДокумент2 страницыBiopsia de Piel1718h9798sОценок пока нет
- SNC-Vía PiramidalДокумент10 страницSNC-Vía Piramidal1718h9798sОценок пока нет
- Historia de La Medicina Legal en EcuadorДокумент11 страницHistoria de La Medicina Legal en Ecuador1718h9798sОценок пока нет
- Plan de NegociosДокумент42 страницыPlan de Negocios1718h9798sОценок пока нет
- Caso Practico BroncodilatadoresДокумент9 страницCaso Practico Broncodilatadores1718h9798sОценок пока нет
- Paresia - Parálisis - Síntomas de La Vía PiramidalДокумент4 страницыParesia - Parálisis - Síntomas de La Vía Piramidal1718h9798sОценок пока нет
- Técnica de La Frecuencia CardiacaДокумент2 страницыTécnica de La Frecuencia Cardiaca1718h9798sОценок пока нет
- Caso Clínico Anafilaxia AsmaДокумент2 страницыCaso Clínico Anafilaxia Asma1718h9798sОценок пока нет
- Auscultacion PulmonarДокумент32 страницыAuscultacion Pulmonar1718h9798sОценок пока нет
- Translocación BacterianaДокумент3 страницыTranslocación Bacteriana1718h9798sОценок пока нет
- Preguntas InmunologiaДокумент9 страницPreguntas Inmunologia1718h9798sОценок пока нет
- ExploracionfisicaДокумент14 страницExploracionfisica1718h9798sОценок пока нет
- Lanzamientos y SaltosДокумент30 страницLanzamientos y Saltos1718h9798sОценок пока нет
- EL PULSO GuarderasДокумент2 страницыEL PULSO Guarderas1718h9798sОценок пока нет
- Tronco BraquiocefálicoДокумент6 страницTronco Braquiocefálico1718h9798sОценок пока нет
- Desarrollo NeoplasiaДокумент4 страницыDesarrollo Neoplasia1718h9798sОценок пока нет
- Conclusiones VitaminasДокумент3 страницыConclusiones Vitaminas1718h9798sОценок пока нет
- Mecanismos de Lesión CelularДокумент4 страницыMecanismos de Lesión Celular1718h9798sОценок пока нет
- Curso de Bioenergetica-Módulo 2Документ22 страницыCurso de Bioenergetica-Módulo 2Roberto Ortega93% (15)
- Aparato DigestivoДокумент18 страницAparato DigestivoVane Jth100% (1)
- Pacto LeoninoДокумент6 страницPacto LeoninoCelcar TravelAgencyОценок пока нет
- CIPRESДокумент2 страницыCIPRESFlor LopezОценок пока нет
- 14 - Manual Cto - OftalmologiaДокумент81 страница14 - Manual Cto - OftalmologiaVictor CutiОценок пока нет
- 10 ChistesДокумент12 страниц10 ChistesJesús GarcíaОценок пока нет
- Craneados, VertebradosДокумент3 страницыCraneados, VertebradosMel GómezОценок пока нет
- La PlacentaДокумент16 страницLa PlacentaDiego Rengifo ValenciaОценок пока нет
- Examen Fisico NeonatoДокумент9 страницExamen Fisico NeonatoLorenaBernalОценок пока нет
- El Cuero - Cartilla 2 (3) Taller3Документ20 страницEl Cuero - Cartilla 2 (3) Taller3Cópernico GalileiОценок пока нет
- Monografía Del CuentoДокумент15 страницMonografía Del CuentoMélany Solansh MeléndezОценок пока нет
- Danza Chuchulayas PDFДокумент6 страницDanza Chuchulayas PDFMarco TorresОценок пока нет
- 1 HematopoyesisДокумент35 страниц1 HematopoyesisRichard Gonzales DíazОценок пока нет
- Anatomia 2013 - Semana 2Документ4 страницыAnatomia 2013 - Semana 2Beltran Amaro Bravo ChávezОценок пока нет
- ¿Por Qué Late El Corazón?: Tesoro de La JuventudДокумент2 страницы¿Por Qué Late El Corazón?: Tesoro de La JuventudKalem SamiОценок пока нет
- Deber 4Документ1 страницаDeber 4Brayan CarreraОценок пока нет
- T13 Reacciones PDFДокумент17 страницT13 Reacciones PDFcarlos esteban diaz aragonОценок пока нет
- Mecano ReceptoresДокумент9 страницMecano ReceptoresRaul AntonioОценок пока нет
- Polita Va A La EscuelaДокумент69 страницPolita Va A La EscuelaCarolina Olavarria88% (8)
- Taller de RelajacionДокумент18 страницTaller de RelajacionbezalithОценок пока нет
- Criterios de ROMA III PDFДокумент12 страницCriterios de ROMA III PDFLuis Miguel Acuña López100% (1)
- Descripción Del Aparato Reproductor Masculino y Femenino Del Cavia PorcellusДокумент6 страницDescripción Del Aparato Reproductor Masculino y Femenino Del Cavia PorcellusNatalia Hernandez UribeОценок пока нет
- AnatomíaДокумент17 страницAnatomíaIng Arturo Rodríguez SerquénОценок пока нет
- Cria OVINOS Sist Semi-EstabДокумент21 страницаCria OVINOS Sist Semi-EstabManuel Ruiz Zebadua100% (2)
- Anécdotas y Resumen de Los Inicios Taurinos de "EL VITI": Santiago MartínДокумент20 страницAnécdotas y Resumen de Los Inicios Taurinos de "EL VITI": Santiago MartíntourismaldeadavilaОценок пока нет
- Historia ClinicaДокумент40 страницHistoria ClinicaFercha NaranjoОценок пока нет
- Resumen de InflamaciónДокумент9 страницResumen de InflamaciónAleyeli CordovaОценок пока нет
- Frenillos PDFДокумент77 страницFrenillos PDFCarolina Diaz100% (1)
- Informe UvaДокумент31 страницаInforme UvaHoracio QH100% (1)
- Arnés o Collar para Tu PerroДокумент5 страницArnés o Collar para Tu PerroGonzalo SucaОценок пока нет