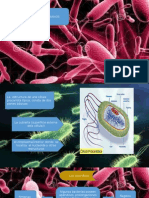
Вам также может понравиться
- Flagelo BacteriaanoДокумент35 страницFlagelo Bacteriaanol3anng100% (1)
- Hemidesmosomas y DesmosomasДокумент3 страницыHemidesmosomas y Desmosomasjimmy belloОценок пока нет
- Lab 6 GeneticaДокумент7 страницLab 6 GeneticaLynda HïcklïnОценок пока нет
- 5 Flagelo y PiliДокумент28 страниц5 Flagelo y PiliAmari SolisОценок пока нет
- Apendices BacterianosДокумент11 страницApendices BacterianosAndrea PilarОценок пока нет
- Flagelos BacterianosДокумент12 страницFlagelos BacterianosRicardo SandovalОценок пока нет
- Flagelo BacterianoДокумент10 страницFlagelo BacterianoKENIAОценок пока нет
- Preguntas de Repaso MicrobiologiaДокумент5 страницPreguntas de Repaso MicrobiologiaCesarAugustoGamioMaldonadoОценок пока нет
- Bacteria - Actividades Unidad 7Документ12 страницBacteria - Actividades Unidad 7Lizzet SalazarОценок пока нет
- MG TEMA 4movilidad+bacterianaДокумент24 страницыMG TEMA 4movilidad+bacterianaBryan JumboОценок пока нет
- Tema 5Документ8 страницTema 5DIablo88Оценок пока нет
- Flagelos OficialДокумент21 страницаFlagelos OficialMarisol BravoОценок пока нет
- BiologiaДокумент24 страницыBiologiaSegundo ChelaОценок пока нет
- Ensayo 3 MicroДокумент10 страницEnsayo 3 MicroStefania SaltosОценок пока нет
- Movilidad BacterianaДокумент24 страницыMovilidad BacterianaCesar Eduardo Pastrano QuiñaОценок пока нет
- UntitledДокумент31 страницаUntitledraquel quiñonezОценок пока нет
- Bibliografia Cel Procarioticas y Eucarioticas Clase 2Документ8 страницBibliografia Cel Procarioticas y Eucarioticas Clase 2Nancy BertucciОценок пока нет
- Elementos FacultativosДокумент8 страницElementos FacultativosVILMA HUANCAHUARI BEDRIÑANAОценок пока нет
- Generalidades Bacterio Unidad 1Документ15 страницGeneralidades Bacterio Unidad 1Maria Virginia DzysiukОценок пока нет
- Citoplasma BacterianoДокумент17 страницCitoplasma Bacterianol3anngОценок пока нет
- Bio. (08) Celula Procariota 53 - 60Документ8 страницBio. (08) Celula Procariota 53 - 60ALEXANDER DAVID LOPEZ ESCOBEDOОценок пока нет
- La Celula - LlojaaaaaaaaaaaДокумент112 страницLa Celula - LlojaaaaaaaaaaasoldjairОценок пока нет
- Biologia - Seminario 6 )Документ12 страницBiologia - Seminario 6 )karla_c_94Оценок пока нет
- .X Documento 1 MICROBIOLOGIA CLINICAДокумент30 страниц.X Documento 1 MICROBIOLOGIA CLINICAAnamaria AlvarezОценок пока нет
- C Loro PlastoДокумент11 страницC Loro PlastoYuly EstefyОценок пока нет
- Microbiología U4Документ10 страницMicrobiología U4Hansony MarОценок пока нет
- Tema 27 12Документ35 страницTema 27 12MaríaОценок пока нет
- FlagelosДокумент4 страницыFlagelosJorge RamirezОценок пока нет
- TEMA 8 Membrana Plasmatica y Organulos MembranososДокумент22 страницыTEMA 8 Membrana Plasmatica y Organulos MembranososARIADNA MÉNDEZ ANCHUELOОценок пока нет
- Complemento Teórico Célula VegetalДокумент13 страницComplemento Teórico Célula VegetalIlse Legaria AguilarОценок пока нет
- U2Документ144 страницыU2Matías BacherОценок пока нет
- Estructura de La Membrana Celular y Modelo MosaicoДокумент10 страницEstructura de La Membrana Celular y Modelo MosaicoRUBI VAZQUEZОценок пока нет
- Células y TejidosДокумент17 страницCélulas y TejidospedroОценок пока нет
- Microbiologia Capitulo 3Документ4 страницыMicrobiologia Capitulo 3Stéfano Cirigliano PodestáОценок пока нет
- Flagelos BacterianosДокумент12 страницFlagelos BacterianosDarwinA.MillaОценок пока нет
- Librito 2,3,4 y 6Документ23 страницыLibrito 2,3,4 y 6Nicole Antonella RiosОценок пока нет
- Tarea 2 ORGANIZACION DE LA CELULAДокумент14 страницTarea 2 ORGANIZACION DE LA CELULASantiago GiraldoОценок пока нет
- CITOLOGÍAДокумент80 страницCITOLOGÍAKetty Roxana GTerronesОценок пока нет
- UBA - CBC - Biologia - Resumen Libritos 2, 3, 4 y 6 - CatДокумент18 страницUBA - CBC - Biologia - Resumen Libritos 2, 3, 4 y 6 - CatJuanJoseОценок пока нет
- EXPERIMENTO 7 Cilios y FlagelosДокумент8 страницEXPERIMENTO 7 Cilios y FlagelosLizsavelSV100% (1)
- Ultraestructura Del FlageloДокумент2 страницыUltraestructura Del FlageloClarita GallardoОценок пока нет
- Flagelo BacterianoДокумент3 страницыFlagelo BacterianoSusy SaenzОценок пока нет
- Tema 29Документ20 страницTema 29Antonio IllanaОценок пока нет
- Vesicle DockngДокумент20 страницVesicle DockngPamela AlarcónОценок пока нет
- MicroooooooДокумент8 страницMicroooooooSheyla CarrascalОценок пока нет
- Clase 2Документ11 страницClase 2Joss AlvaradoОценок пока нет
- 9D - Sem 08 - Biologia - PRДокумент8 страниц9D - Sem 08 - Biologia - PRJorge escobedoОценок пока нет
- Organulo Celular IДокумент74 страницыOrganulo Celular IRaikmg martinОценок пока нет
- Guia 4 Celula y Organelos Bacterianos y BiofilmДокумент5 страницGuia 4 Celula y Organelos Bacterianos y BiofilmVerónica GranadosОценок пока нет
- Resumen Biología Examen 18 - 01Документ13 страницResumen Biología Examen 18 - 01adnОценок пока нет
- Celula ProcaritoaДокумент34 страницыCelula Procaritoajuan carlosОценок пока нет
- La CelulaДокумент18 страницLa CelulavivitordeОценок пока нет
- Clase 2 - Morfología BacterianaДокумент88 страницClase 2 - Morfología BacterianaMiguel Alejandro Moreno Pitti100% (1)
- 20170908220928 (1)Документ41 страница20170908220928 (1)Evelin Ramos MarcelianoОценок пока нет
- FlagelosДокумент6 страницFlagelosRaul BasilioОценок пока нет
- Preinforme Células Vegetales e InclusionesДокумент9 страницPreinforme Células Vegetales e InclusionesDavid Vinson100% (2)
- Resumen MicroДокумент36 страницResumen MicroNico LopezОценок пока нет
- MICROBIOLOGIAДокумент21 страницаMICROBIOLOGIAAxel SanHerОценок пока нет
- Eucariota VS PROCARIOTAДокумент36 страницEucariota VS PROCARIOTADanny GarayОценок пока нет
- Tema 4 - Organoides CitoplasmaticosДокумент10 страницTema 4 - Organoides CitoplasmaticosYahn Carlos M MОценок пока нет
- Análisis funcional y microbiológico de derivados lácteos y cárnicosОт EverandAnálisis funcional y microbiológico de derivados lácteos y cárnicosОценок пока нет
- Introducción a la Biología: RESÚMENES UNIVERSITARIOSОт EverandIntroducción a la Biología: RESÚMENES UNIVERSITARIOSРейтинг: 5 из 5 звезд5/5 (1)
- Tincion Gram AlumnosДокумент9 страницTincion Gram AlumnosHin UzumakiОценок пока нет
- BiotecnologiaДокумент29 страницBiotecnologiaJaqueline OsorioОценок пока нет
- Enzimas Info.Документ19 страницEnzimas Info.Rossman MejicanosОценок пока нет
- Biocell Junqueira9 Introduccion Formas Celulares Acelulares 2012brrДокумент12 страницBiocell Junqueira9 Introduccion Formas Celulares Acelulares 2012brrGabriel QuimbayoОценок пока нет
- Delitos de Manipulación GenéticaДокумент9 страницDelitos de Manipulación GenéticaCamila VacaОценок пока нет
- Actividad de Aprendizaje CelulasДокумент3 страницыActividad de Aprendizaje CelulasMaycol Callan HernandezОценок пока нет
- Factores Que Influyen en La Penetración de Productos.Документ11 страницFactores Que Influyen en La Penetración de Productos.wilsory giraldoОценок пока нет
- Microbiologia PauДокумент7 страницMicrobiologia PauJoselo TorresОценок пока нет
- Preguntas Selectividad Célula-MetabolismoДокумент4 страницыPreguntas Selectividad Célula-Metabolismobioquimchivite2094100% (2)
- Identificación de La Guía de Aprendizaje 2Документ9 страницIdentificación de La Guía de Aprendizaje 2luz elena rojasОценок пока нет
- Cuadro Comparativo. Estructura CelularДокумент18 страницCuadro Comparativo. Estructura CelularDayana Tonantzin Gómora Esquivel100% (1)
- Infome BioДокумент16 страницInfome BioAnonymous R8mRx1IDKОценок пока нет
- Bio SelectividadДокумент32 страницыBio SelectividadcОценок пока нет
- Actividades 1 y 2 MembranaДокумент2 страницыActividades 1 y 2 MembranaIngridAmayaCarrascoОценок пока нет
- Manual de Biología 2022 Primer ParcialДокумент56 страницManual de Biología 2022 Primer Parcialpapeleria ciberborealОценок пока нет
- Malla Curricular de Ciencias Naturales Segundo BasicoДокумент15 страницMalla Curricular de Ciencias Naturales Segundo BasicoElan Mendoza100% (2)
- Carpeta Micro PDFДокумент93 страницыCarpeta Micro PDFjustinaОценок пока нет
- Primer Bloque de PreguntasДокумент26 страницPrimer Bloque de Preguntasmr.wilblackОценок пока нет
- Sistema CelularДокумент31 страницаSistema CelularDiana Hernandez100% (1)
- Ciclo Del FosforoДокумент6 страницCiclo Del FosforoffffОценок пока нет
- Pruebas Tipo Icfes SextoДокумент6 страницPruebas Tipo Icfes SextoHarold HajoarОценок пока нет
- Ficha #1 - Seres VivosДокумент8 страницFicha #1 - Seres VivosSusan LinaresОценок пока нет
- Radioterapia Ley de Bergonie y TribondeauДокумент3 страницыRadioterapia Ley de Bergonie y TribondeauCastañeda Reyna100% (1)
- Merkaba 4Документ15 страницMerkaba 4lector12Оценок пока нет
- YayaДокумент3 страницыYayaeliuseduardo sagastumemoratyaОценок пока нет
- Célula, Tejido ResumenДокумент85 страницCélula, Tejido ResumenJuan RamónОценок пока нет
- EvaluaciónДокумент1 страницаEvaluaciónMaria Fernanda Cortizas UrquijoОценок пока нет
- Discovery: PruebaДокумент28 страницDiscovery: PruebaDorys López COAIPYME RLОценок пока нет