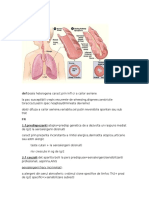
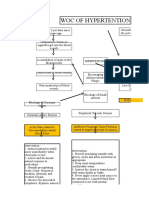
Вам также может понравиться
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeОт EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeРейтинг: 4 из 5 звезд4/5 (5794)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreОт EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreРейтинг: 4 из 5 звезд4/5 (1090)
- Never Split the Difference: Negotiating As If Your Life Depended On ItОт EverandNever Split the Difference: Negotiating As If Your Life Depended On ItРейтинг: 4.5 из 5 звезд4.5/5 (838)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceОт EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceРейтинг: 4 из 5 звезд4/5 (895)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersОт EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersРейтинг: 4.5 из 5 звезд4.5/5 (344)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureОт EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureРейтинг: 4.5 из 5 звезд4.5/5 (474)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)От EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Рейтинг: 4.5 из 5 звезд4.5/5 (121)
- The Emperor of All Maladies: A Biography of CancerОт EverandThe Emperor of All Maladies: A Biography of CancerРейтинг: 4.5 из 5 звезд4.5/5 (271)
- The Little Book of Hygge: Danish Secrets to Happy LivingОт EverandThe Little Book of Hygge: Danish Secrets to Happy LivingРейтинг: 3.5 из 5 звезд3.5/5 (400)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyОт EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyРейтинг: 3.5 из 5 звезд3.5/5 (2259)
- The Yellow House: A Memoir (2019 National Book Award Winner)От EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Рейтинг: 4 из 5 звезд4/5 (98)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaОт EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaРейтинг: 4.5 из 5 звезд4.5/5 (266)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryОт EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryРейтинг: 3.5 из 5 звезд3.5/5 (231)
- Team of Rivals: The Political Genius of Abraham LincolnОт EverandTeam of Rivals: The Political Genius of Abraham LincolnРейтинг: 4.5 из 5 звезд4.5/5 (234)
- The Unwinding: An Inner History of the New AmericaОт EverandThe Unwinding: An Inner History of the New AmericaРейтинг: 4 из 5 звезд4/5 (45)
- Light and The Human Biofield: Dietrich KlinghardtДокумент13 страницLight and The Human Biofield: Dietrich KlinghardtframistatОценок пока нет
- Coduri DiagnosticeДокумент3 страницыCoduri DiagnosticeIrina AdrianaОценок пока нет
- Radiographic Anatomy PDFДокумент314 страницRadiographic Anatomy PDFIrina Adriana0% (1)
- Chest: Advancing Respiratory ResearchДокумент5 страницChest: Advancing Respiratory ResearchIrina AdrianaОценок пока нет
- Sleep Apnea Research in AnimalsДокумент20 страницSleep Apnea Research in AnimalsIrina AdrianaОценок пока нет
- Wms Main Pocket Guide 2017Документ29 страницWms Main Pocket Guide 2017justrudinОценок пока нет
- Quantitative Electroencephalogram Measures in Adult Obstructive Sleep Apnea - Potential Biomarkers of Neurobehavioural Functioning - ScienceDirectДокумент50 страницQuantitative Electroencephalogram Measures in Adult Obstructive Sleep Apnea - Potential Biomarkers of Neurobehavioural Functioning - ScienceDirectIrina AdrianaОценок пока нет
- Sleep Apnea Research in AnimalsДокумент20 страницSleep Apnea Research in AnimalsIrina AdrianaОценок пока нет
- Coduri DiagnosticeДокумент3 страницыCoduri DiagnosticeIrina AdrianaОценок пока нет
- Animal Research Sleep ApneaДокумент8 страницAnimal Research Sleep ApneaIrina AdrianaОценок пока нет
- Wms GOLD 2017 Pocket Guide 1 PDFДокумент42 страницыWms GOLD 2017 Pocket Guide 1 PDFL P PutriОценок пока нет
- Coduri DiagnosticeДокумент3 страницыCoduri DiagnosticeIrina AdrianaОценок пока нет
- Diabetul GestationalДокумент6 страницDiabetul GestationalIrina AdrianaОценок пока нет
- Diagnostic SarcinaДокумент3 страницыDiagnostic SarcinaIrina AdrianaОценок пока нет
- Medicatie CardiologieДокумент6 страницMedicatie CardiologieIrina AdrianaОценок пока нет
- Examinarea Clinica in ObstetricaДокумент12 страницExaminarea Clinica in ObstetricaDiana AndreiОценок пока нет
- Nasterea in Prezentatie PelvinaДокумент14 страницNasterea in Prezentatie PelvinaIrina AdrianaОценок пока нет
- AstmДокумент10 страницAstmIrina AdrianaОценок пока нет
- Diagnostic SarcinaДокумент3 страницыDiagnostic SarcinaIrina AdrianaОценок пока нет
- Vaginala Vs Cezariana-Articol RevistaДокумент10 страницVaginala Vs Cezariana-Articol RevistaIrina AdrianaОценок пока нет
- F Abrevieri, Bibl, CopДокумент8 страницF Abrevieri, Bibl, CopAdelina PapanceaОценок пока нет
- Vasoconstriction in Septic Shock: What'S New in Intensive CareДокумент4 страницыVasoconstriction in Septic Shock: What'S New in Intensive CareGabriela PachecoОценок пока нет
- Renal Blood Flow Until Regulation of RBF and GFRДокумент5 страницRenal Blood Flow Until Regulation of RBF and GFRDoogie ReynaldoОценок пока нет
- Use of Vasopressors and InotropesДокумент16 страницUse of Vasopressors and InotropesIrving H Torres LopezОценок пока нет
- SympatheticДокумент10 страницSympatheticSharneeshriyaОценок пока нет
- Effects of HypoxiaДокумент6 страницEffects of HypoxiaMaria TelloОценок пока нет
- Public Assessment Report Decentralised ProcedureДокумент26 страницPublic Assessment Report Decentralised Procedurefad12345Оценок пока нет
- Opie 1997Документ9 страницOpie 1997Arbi SulaimanОценок пока нет
- ErgotamineДокумент2 страницыErgotamineAnonymous xv9UDvKLОценок пока нет
- Ciclo Nasal RespiratorioДокумент7 страницCiclo Nasal RespiratorioManuel RaGalОценок пока нет
- Makalah Bahasa Inggris Stik AvicennaДокумент16 страницMakalah Bahasa Inggris Stik AvicennahemraninОценок пока нет
- Heat and Cold Therapy PPT 17Документ54 страницыHeat and Cold Therapy PPT 17sargunavalli balrajОценок пока нет
- WOC CaseДокумент4 страницыWOC Casedwi adindaОценок пока нет
- Should Norepinephrine, Rather Than PhenylephrineДокумент8 страницShould Norepinephrine, Rather Than PhenylephrineDiego Andres Rojas TejadaОценок пока нет
- Sean Walsh, Emma King - Pulse Diagnosis - A Clinical Guide, 1e-Churchill Livingstone (2007) PDFДокумент427 страницSean Walsh, Emma King - Pulse Diagnosis - A Clinical Guide, 1e-Churchill Livingstone (2007) PDFLM_S_S100% (1)
- Phenylephrine: Pilar D'Ocon, Universidad de Valencia, Valencia, SpainДокумент6 страницPhenylephrine: Pilar D'Ocon, Universidad de Valencia, Valencia, Spainarief budiman1Оценок пока нет
- 5.2 Human Nervous System 2 MSДокумент9 страниц5.2 Human Nervous System 2 MSTosyn AriyoОценок пока нет
- Local Anesthesia Part-1Документ33 страницыLocal Anesthesia Part-1Abdul RazakОценок пока нет
- BP602T Unit 1-3Документ207 страницBP602T Unit 1-3Gyampoh SolomonОценок пока нет
- Vasoconstrictors - Indications and PrecautionsДокумент14 страницVasoconstrictors - Indications and PrecautionsJUAREZ GONZALEZ EDWARD HASSANОценок пока нет
- Hans SelyeДокумент10 страницHans SelyeZeyrra Mariana100% (2)
- @anesthesia - Books 2015 MCQs For Handbook of Local Anesthesia PDFДокумент205 страниц@anesthesia - Books 2015 MCQs For Handbook of Local Anesthesia PDFVivek Sinha100% (2)
- Fisiologia Do Sistema Cardiovascular II e III - TextoДокумент6 страницFisiologia Do Sistema Cardiovascular II e III - TextoDavidson De AraújoОценок пока нет
- Sinarest AF Syr Drops2018Документ4 страницыSinarest AF Syr Drops2018Priya EaswarОценок пока нет
- Serotonin PPT Notes 2023Документ20 страницSerotonin PPT Notes 2023adhi85099Оценок пока нет
- Cafergot: Name of The MedicineДокумент10 страницCafergot: Name of The MedicinePoom ColicОценок пока нет
- Adrenergic DrugsДокумент2 страницыAdrenergic DrugsSunshine_Bacla_4275Оценок пока нет
- Step 1 Physiology Flash CardsДокумент22 страницыStep 1 Physiology Flash CardsbobiomeОценок пока нет
- The Circulation: Blood Pressure RegulationДокумент42 страницыThe Circulation: Blood Pressure RegulationLouis JinОценок пока нет
- PCOL CHP 4-6 (Situational Analysis)Документ2 страницыPCOL CHP 4-6 (Situational Analysis)Sunshine_Bacla_4275Оценок пока нет