Вам также может понравиться
- Rampa, Lobsang - El Tercer Ojo PDFДокумент187 страницRampa, Lobsang - El Tercer Ojo PDFAni Yamamoto Akiba100% (1)
- Ciclo de Krebs Seminario 2dauДокумент8 страницCiclo de Krebs Seminario 2dauDanixaKristelBarriosUcañayОценок пока нет
- Calculadora de TannerДокумент8 страницCalculadora de TannerFootball Club Allianz100% (1)
- Yoga Arte y Ciencia de Vida-Indra DeviДокумент16 страницYoga Arte y Ciencia de Vida-Indra DeviMilagros García100% (3)
- DR 3900 Manual HACHДокумент150 страницDR 3900 Manual HACHAngeles AvilaОценок пока нет
- Pets SoldaduraДокумент2 страницыPets SoldaduraJulioMedina100% (4)
- Ciclo de KrebsДокумент7 страницCiclo de KrebsFranciscoSalvadorОценок пока нет
- Mapa Conceptual de Carbohidratos y Ensayo Del Ciclo de KrebsДокумент17 страницMapa Conceptual de Carbohidratos y Ensayo Del Ciclo de KrebsJuan EnseñatОценок пока нет
- Ciclo de KrebsДокумент9 страницCiclo de KrebsJohnHarrisonОценок пока нет
- Resumen 12 Ciclo de KrebsДокумент12 страницResumen 12 Ciclo de KrebsMariam Cabellos SОценок пока нет
- Ciclo Del Ácido Cítrico y GlucolisisДокумент18 страницCiclo Del Ácido Cítrico y GlucolisisJerry Carhuatanta SaavedraОценок пока нет
- Resumen de Rutas Metabólicas BCMДокумент52 страницыResumen de Rutas Metabólicas BCMLuana HuertasОценок пока нет
- Ciclo de KrebsДокумент22 страницыCiclo de KrebsReben Zecnaj100% (1)
- Tema 4. Acetil-CoA y Ciclo de Krebs.Документ14 страницTema 4. Acetil-CoA y Ciclo de Krebs.Manuel Marin DominguezОценок пока нет
- Ciclo de KrebsДокумент19 страницCiclo de KrebsWikii Wikoso0% (1)
- RESPIRACION CELULAR-Valeryn Valera-12Документ5 страницRESPIRACION CELULAR-Valeryn Valera-12Valeryn Valera JiménezОценок пока нет
- Rutas MetabólicasДокумент9 страницRutas MetabólicasSelene De León Morga100% (4)
- Ciclo de KrebsДокумент9 страницCiclo de KrebsMileОценок пока нет
- CicloK GeorgetteДокумент6 страницCicloK Georgetteosmigdy mataОценок пока нет
- Ciclo de KrebsДокумент33 страницыCiclo de KrebsLN Brenda Janeth Beltrán DíazОценок пока нет
- LipogénesisДокумент16 страницLipogénesisgabycita910% (1)
- Ciclo de KrebsДокумент10 страницCiclo de KrebsMariana crystel Cabanillas SalcidoОценок пока нет
- NUTRICIONДокумент6 страницNUTRICIONEl anonimo oscuroОценок пока нет
- Ciclo Del Ácido CítricoДокумент5 страницCiclo Del Ácido CítricoGENESIS HERNANDEZОценок пока нет
- Ciclo de KrebsДокумент14 страницCiclo de KrebsJhonatan AguilarОценок пока нет
- Procesos Oxidativos 1Документ71 страницаProcesos Oxidativos 1alexander naspiranОценок пока нет
- El CICLO de KREBS y Su RegulaciónДокумент9 страницEl CICLO de KREBS y Su Regulaciónluis emilio pachecoОценок пока нет
- Ciclo de Krebs 22Документ11 страницCiclo de Krebs 22Mari CastedoОценок пока нет
- Ciclo de KrebsДокумент21 страницаCiclo de KrebsAndresОценок пока нет
- Ciclo de KlebsДокумент9 страницCiclo de KlebsIram Fernando Oleta CruzОценок пока нет
- Ciclo de Krebs - BiologiaДокумент6 страницCiclo de Krebs - BiologiaCARLA JOMBIОценок пока нет
- Ashley Salcedo. 100478923. Sec 04. Respiracion CelularДокумент5 страницAshley Salcedo. 100478923. Sec 04. Respiracion CelularLenny MotaОценок пока нет
- Objetivos Metabolismo y Ciclo de Krebs, Metabolismo de CarbohidratosДокумент23 страницыObjetivos Metabolismo y Ciclo de Krebs, Metabolismo de Carbohidratosapi-369665690% (10)
- Ciclo de KrebsДокумент12 страницCiclo de KrebsGallardo RichardОценок пока нет
- Colegio Adventista PimentelДокумент26 страницColegio Adventista PimentelCamila RubioОценок пока нет
- ESQUEMA DE METABOLISMOS y CICLO DE KREBSДокумент3 страницыESQUEMA DE METABOLISMOS y CICLO DE KREBSJavier Hector Escobari LimpiasОценок пока нет
- Ciclo de KrebsДокумент8 страницCiclo de KrebsGustavo Adolfo MozОценок пока нет
- Bioquimica MetabolicaДокумент15 страницBioquimica MetabolicaNubia Patricia Parada RiveraОценок пока нет
- Ciclo de KrebsДокумент6 страницCiclo de KrebsJose Luis Arroyo Hernandez0% (1)
- CICLO DE KREBS Power PointДокумент19 страницCICLO DE KREBS Power PointAnibal Munguia67% (3)
- Tema 14 Ciclo de KrebsДокумент6 страницTema 14 Ciclo de KrebsValentinaОценок пока нет
- Ciclo de KrebsДокумент58 страницCiclo de Krebssantos ArchivaldoОценок пока нет
- Artículo Del Ciclo de Krebs Mayo 31 2018Документ5 страницArtículo Del Ciclo de Krebs Mayo 31 2018Lesdy MarllyОценок пока нет
- Ciclo de KrebsДокумент13 страницCiclo de KrebsCRISTHIAN GONZALES CATAYОценок пока нет
- Metabolismo INGДокумент16 страницMetabolismo INGLuis Felipe Diaz GonzalezОценок пока нет
- Función Del Ciclo Krebs en El MetabolismoДокумент8 страницFunción Del Ciclo Krebs en El MetabolismoJorge Alvaro Mendez MolinaОценок пока нет
- Bioquimica 3Документ14 страницBioquimica 3adonayОценок пока нет
- Metabolismo y Respiracion CelularДокумент25 страницMetabolismo y Respiracion CelularValeria GuerraОценок пока нет
- 2ciclo de KrebsДокумент6 страниц2ciclo de KrebsSanti GarciaОценок пока нет
- Ciclo de Krebs y Vía AnapleróticasДокумент7 страницCiclo de Krebs y Vía AnapleróticasDelgado Mejía BrandonОценок пока нет
- Metabolismo Leidy Tatiana AcevedoДокумент16 страницMetabolismo Leidy Tatiana AcevedoÄcëvëdö TatianaОценок пока нет
- Laura BioquimicaДокумент19 страницLaura BioquimicaLauren AmayaОценок пока нет
- Ciclo de KrebsДокумент33 страницыCiclo de KrebsJosé Álvarez RoblesОценок пока нет
- Acetil-CoA, Ciclo de KrebsДокумент12 страницAcetil-CoA, Ciclo de KrebsClauMcClauОценок пока нет
- Ciclo de Krebs y Glucolisis - Equipo 7 - 6HДокумент20 страницCiclo de Krebs y Glucolisis - Equipo 7 - 6HEmiliano LoAlОценок пока нет
- Ciclo de Krebs 2019Документ36 страницCiclo de Krebs 2019KarlaRamosОценок пока нет
- Ciclo de Krebs, Cadena Respiratoria y Fosforilación OxidativaДокумент8 страницCiclo de Krebs, Cadena Respiratoria y Fosforilación OxidativaJohan Fernando HerreraОценок пока нет
- Tarea Creativa IdeasДокумент2 страницыTarea Creativa IdeasjuanmideluxeОценок пока нет
- CICLO DE KREBS y GlucolisisДокумент7 страницCICLO DE KREBS y GlucolisisRodrigo MirandaОценок пока нет
- Bioquímica 18 16 II.UДокумент29 страницBioquímica 18 16 II.ULorena LisbethОценок пока нет
- Regulación EnzimáticaДокумент8 страницRegulación Enzimáticaclaudia karina almaguer hidalgoОценок пока нет
- CUESTIONARIOДокумент8 страницCUESTIONARIOJuan Jose Baros AyalaОценок пока нет
- Ciclo Del Acido Citrico y Reacciones AnapleroticasДокумент5 страницCiclo Del Acido Citrico y Reacciones AnapleroticasLALUKISОценок пока нет
- La Ciencia Del Metabolismo: Metabolismo sin rodeosОт EverandLa Ciencia Del Metabolismo: Metabolismo sin rodeosРейтинг: 5 из 5 звезд5/5 (1)
- Estadios de Taner EtapasДокумент2 страницыEstadios de Taner EtapasGregorio ChavezОценок пока нет
- Cómo Dejar Las Drogas: Sin Síndrome de AbstinenciaДокумент6 страницCómo Dejar Las Drogas: Sin Síndrome de AbstinenciaFootball Club AllianzОценок пока нет
- 12 Desarrollo SexualДокумент5 страниц12 Desarrollo SexualFootball Club AllianzОценок пока нет
- Ciclo de KrebsДокумент9 страницCiclo de KrebsFootball Club AllianzОценок пока нет
- El Sexo Adolescente Es CotidianoДокумент5 страницEl Sexo Adolescente Es CotidianoFootball Club AllianzОценок пока нет
- Cómo Poner Inyecciones CorrectamenteДокумент5 страницCómo Poner Inyecciones CorrectamenteFootball Club AllianzОценок пока нет
- Desarrollo Psicosexual FreudДокумент24 страницыDesarrollo Psicosexual FreudSonido SurrealОценок пока нет
- La Tutela de Los Menores de EdadДокумент2 страницыLa Tutela de Los Menores de EdadFootball Club AllianzОценок пока нет
- Rendimiento Deportivo PaidotrivoДокумент10 страницRendimiento Deportivo PaidotrivoMauricio Alexander Ortiz GamboaОценок пока нет
- CHINEA CARLOS S - Pensamiento Lateral Y AcertijosДокумент12 страницCHINEA CARLOS S - Pensamiento Lateral Y AcertijosFootball Club AllianzОценок пока нет
- Cómo Rezar El Santo Rosario CatólicoДокумент3 страницыCómo Rezar El Santo Rosario Católicovaticanocatolico.com100% (9)
- CursoISAK PDFДокумент9 страницCursoISAK PDFFootball Club AllianzОценок пока нет
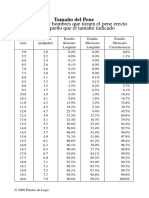
- Tamano NormalДокумент3 страницыTamano NormalAarón MontoyaОценок пока нет
- 3606 - Especial Agentes de Jugadores AFAДокумент8 страниц3606 - Especial Agentes de Jugadores AFAFootball Club AllianzОценок пока нет
- Apuntes de Matematicas Basicas (Algebra, Calculo Etc)Документ117 страницApuntes de Matematicas Basicas (Algebra, Calculo Etc)Gabriel MenchuОценок пока нет
- Mate Ma Tic A Basic A 1Документ139 страницMate Ma Tic A Basic A 1Rhandorl TejadaОценок пока нет
- Así Pensaba Markus - Capitulo 1Документ2 страницыAsí Pensaba Markus - Capitulo 1Football Club AllianzОценок пока нет
- ANTROPOMETRÍAДокумент21 страницаANTROPOMETRÍAEvelyn Stella Pacheco ArzuzaОценок пока нет
- Evaluación Antropométrica en Niños de 6 A 12 Años de La Ciudad de Corrientes en El Año 2004Документ4 страницыEvaluación Antropométrica en Niños de 6 A 12 Años de La Ciudad de Corrientes en El Año 2004Football Club AllianzОценок пока нет
- Gaete PDFДокумент41 страницаGaete PDFFootball Club Allianz100% (1)
- Sábado CalentitoДокумент3 страницыSábado CalentitoFootball Club AllianzОценок пока нет
- Antropometriayenifer 110902092300 Phpapp02Документ11 страницAntropometriayenifer 110902092300 Phpapp02Football Club AllianzОценок пока нет
- Caracterización Del Entrenador de Alto RendimientoДокумент23 страницыCaracterización Del Entrenador de Alto RendimientoBoris Favian ReyesОценок пока нет
- CV Profe Nicolascaputo 2018Документ2 страницыCV Profe Nicolascaputo 2018Football Club AllianzОценок пока нет
- Curriculum Vitae GECДокумент2 страницыCurriculum Vitae GECFootball Club AllianzОценок пока нет
- Lo Que No Es Una Escuela de FútbolДокумент4 страницыLo Que No Es Una Escuela de FútbolFootball Club AllianzОценок пока нет
- Libro 1Документ2 страницыLibro 1Football Club AllianzОценок пока нет
- Caracteristicas de La Materia VivaДокумент15 страницCaracteristicas de La Materia VivaAnnete RomeroОценок пока нет
- Fluidos de PerforacionДокумент7 страницFluidos de PerforacionjoschirinoОценок пока нет
- Introduccion Hidrometalurgia (Procesos HidrometalurgicosДокумент75 страницIntroduccion Hidrometalurgia (Procesos Hidrometalurgicosjrotapia2002Оценок пока нет
- Cuadernillo Taller 4° Año - TMT.TДокумент25 страницCuadernillo Taller 4° Año - TMT.TLucas VarelaОценок пока нет
- Factor Z para 2 Fases (Traduccion)Документ4 страницыFactor Z para 2 Fases (Traduccion)Itaty Baños100% (1)
- Metabolismo de Carbohidratos Trabajo Grupal 2020 MBH (Autoguardado)Документ14 страницMetabolismo de Carbohidratos Trabajo Grupal 2020 MBH (Autoguardado)Delgado H FabianОценок пока нет
- Aplicaciones de La Nanoquímica en El Desarrollo de Nuevas Tecnologías: Nano CosméticosДокумент14 страницAplicaciones de La Nanoquímica en El Desarrollo de Nuevas Tecnologías: Nano CosméticospaulinaОценок пока нет
- AditivosДокумент13 страницAditivosHewkardОценок пока нет
- Producto 2Документ20 страницProducto 2Eduardo Vilca calla100% (1)
- Conceptos de Fisico QuimicaДокумент15 страницConceptos de Fisico Quimicaapi-297587367Оценок пока нет
- Foro Calor y TemperaturaДокумент2 страницыForo Calor y TemperaturaMarco Antonio Tasayco Yataco100% (1)
- Obtención de Aceite Esencial BoliviaДокумент16 страницObtención de Aceite Esencial BoliviaRonald Tito Cusiquispe MolleОценок пока нет
- Elizabeth Trabajo Bachiller 2020Документ12 страницElizabeth Trabajo Bachiller 2020ANTERO JOSUE CORDOVA RIOSОценок пока нет
- Presentacion PlasticosДокумент31 страницаPresentacion Plasticoscarlos enrique juarez glez100% (2)
- Sustentacion Trabajo Colaborativo - Escenario 7 - PRIMER BLOQUE-CIENCIAS BASICAS - FLUIDOS Y TERMODINAMICA - (GRUPO B03)Документ4 страницыSustentacion Trabajo Colaborativo - Escenario 7 - PRIMER BLOQUE-CIENCIAS BASICAS - FLUIDOS Y TERMODINAMICA - (GRUPO B03)Alejandro BernalОценок пока нет
- Manual Entrepiso Esp Web 25marzo CompressedДокумент9 страницManual Entrepiso Esp Web 25marzo CompressedromdreОценок пока нет
- 09 Encerado 1. Teoria Del Deslizamiento - Rev 01Документ6 страниц09 Encerado 1. Teoria Del Deslizamiento - Rev 01Carlos MuñizОценок пока нет
- Clase 03 Agregados Grueso y FinoДокумент110 страницClase 03 Agregados Grueso y FinoDorfy Martin Culqui SoplinОценок пока нет
- "Un Mar Que Alumbra": Instituto Thomas JeffersonДокумент16 страниц"Un Mar Que Alumbra": Instituto Thomas JeffersonSaly BОценок пока нет
- Indice PeroxidoДокумент9 страницIndice PeroxidoManuel Guillen ParicelaОценок пока нет
- InformeДокумент13 страницInformeMartin TorresОценок пока нет
- Catabolismo de Los LipidosДокумент16 страницCatabolismo de Los LipidosLuis Ernesto Guzman VelazquezОценок пока нет
- Palas de Aerogenerador Version 28Документ58 страницPalas de Aerogenerador Version 28arturo14051984100% (1)
- AssetsДокумент1 страницаAssetsJesus CurielОценок пока нет
- Carbohidratos Estudiantes PDFДокумент7 страницCarbohidratos Estudiantes PDFLAURA LIZETH LAGUNA TEJADAОценок пока нет
- Tecnologia y Control de Calidad Del AireДокумент6 страницTecnologia y Control de Calidad Del AireAlexia YRPОценок пока нет
- 13 ElectroquimicaДокумент6 страниц13 ElectroquimicaIdowantlove WithmeОценок пока нет
- Indice RiteДокумент7 страницIndice RiteJGabriel LedezmaОценок пока нет