Вам также может понравиться
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryОт EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryРейтинг: 3.5 из 5 звезд3.5/5 (231)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)От EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Рейтинг: 4.5 из 5 звезд4.5/5 (121)
- Never Split the Difference: Negotiating As If Your Life Depended On ItОт EverandNever Split the Difference: Negotiating As If Your Life Depended On ItРейтинг: 4.5 из 5 звезд4.5/5 (838)
- The Little Book of Hygge: Danish Secrets to Happy LivingОт EverandThe Little Book of Hygge: Danish Secrets to Happy LivingРейтинг: 3.5 из 5 звезд3.5/5 (400)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaОт EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaРейтинг: 4.5 из 5 звезд4.5/5 (266)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeОт EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeРейтинг: 4 из 5 звезд4/5 (5794)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreОт EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreРейтинг: 4 из 5 звезд4/5 (1090)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyОт EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyРейтинг: 3.5 из 5 звезд3.5/5 (2259)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersОт EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersРейтинг: 4.5 из 5 звезд4.5/5 (345)
- The Emperor of All Maladies: A Biography of CancerОт EverandThe Emperor of All Maladies: A Biography of CancerРейтинг: 4.5 из 5 звезд4.5/5 (271)
- Team of Rivals: The Political Genius of Abraham LincolnОт EverandTeam of Rivals: The Political Genius of Abraham LincolnРейтинг: 4.5 из 5 звезд4.5/5 (234)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceОт EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceРейтинг: 4 из 5 звезд4/5 (895)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureОт EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureРейтинг: 4.5 из 5 звезд4.5/5 (474)
- Space Hulk - WDДокумент262 страницыSpace Hulk - WDIgor Baranenko100% (1)
- The Yellow House: A Memoir (2019 National Book Award Winner)От EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Рейтинг: 4 из 5 звезд4/5 (98)
- The Unwinding: An Inner History of the New AmericaОт EverandThe Unwinding: An Inner History of the New AmericaРейтинг: 4 из 5 звезд4/5 (45)
- CT SizingДокумент62 страницыCT SizingMohamed TalebОценок пока нет
- CH 1 - Democracy and American PoliticsДокумент9 страницCH 1 - Democracy and American PoliticsAndrew Philip ClarkОценок пока нет
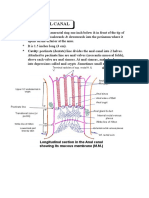
- Anatomy Anal CanalДокумент14 страницAnatomy Anal CanalBela Ronaldoe100% (1)
- Analysis of Electric Machinery Krause Manual Solution PDFДокумент2 страницыAnalysis of Electric Machinery Krause Manual Solution PDFKuldeep25% (8)
- Excon2019 ShowPreview02122019 PDFДокумент492 страницыExcon2019 ShowPreview02122019 PDFSanjay KherОценок пока нет
- Chapter 3 - Organization Structure & CultureДокумент63 страницыChapter 3 - Organization Structure & CultureDr. Shuva GhoshОценок пока нет
- Analizador de Combustion Kigaz 310 Manual EngДокумент60 страницAnalizador de Combustion Kigaz 310 Manual EngJully Milagros Rodriguez LaicheОценок пока нет
- Syllabus DresserДокумент2 страницыSyllabus DresserVikash Aggarwal50% (2)
- 74HC00D 74HC00D 74HC00D 74HC00D: CMOS Digital Integrated Circuits Silicon MonolithicДокумент8 страниц74HC00D 74HC00D 74HC00D 74HC00D: CMOS Digital Integrated Circuits Silicon MonolithicAssistec TecОценок пока нет
- 3 Carbohydrates' StructureДокумент33 страницы3 Carbohydrates' StructureDilan TeodoroОценок пока нет
- NDY 9332v3Документ8 страницNDY 9332v3sulphurdioxideОценок пока нет
- A Case Study Puga Geothermal System,: OF IndiaДокумент7 страницA Case Study Puga Geothermal System,: OF IndiaPERIKALA TARUNОценок пока нет
- Ito Na Talaga Yung FinalДокумент22 страницыIto Na Talaga Yung FinalJonas Gian Miguel MadarangОценок пока нет
- Problem Set-02Документ2 страницыProblem Set-02linn.pa.pa.khaing.2020.2021.fbОценок пока нет
- Recommendations For Students With High Functioning AutismДокумент7 страницRecommendations For Students With High Functioning AutismLucia SaizОценок пока нет
- Unit 16 - Monitoring, Review and Audit by Allan WatsonДокумент29 страницUnit 16 - Monitoring, Review and Audit by Allan WatsonLuqman OsmanОценок пока нет
- Corrosion Fatigue Phenomena Learned From Failure AnalysisДокумент10 страницCorrosion Fatigue Phenomena Learned From Failure AnalysisDavid Jose Velandia MunozОценок пока нет
- Iec TR 61010-3-020-1999Документ76 страницIec TR 61010-3-020-1999Vasko MandilОценок пока нет
- Blue Prism Data Sheet - Provisioning A Blue Prism Database ServerДокумент5 страницBlue Prism Data Sheet - Provisioning A Blue Prism Database Serverreddy_vemula_praveenОценок пока нет
- What Is TranslationДокумент3 страницыWhat Is TranslationSanskriti MehtaОценок пока нет
- AntibioticsДокумент36 страницAntibioticsBen Paolo Cecilia RabaraОценок пока нет
- Lec 33 - Householder MethodДокумент11 страницLec 33 - Householder MethodMudit SinhaОценок пока нет
- CEE Annual Report 2018Документ100 страницCEE Annual Report 2018BusinessTech100% (1)
- SSGC-RSGLEG Draft Study On The Applicability of IAL To Cyber Threats Against Civil AviationДокумент41 страницаSSGC-RSGLEG Draft Study On The Applicability of IAL To Cyber Threats Against Civil AviationPrachita AgrawalОценок пока нет
- On Derridean Différance - UsiefДокумент16 страницOn Derridean Différance - UsiefS JEROME 2070505Оценок пока нет
- E0 UoE Unit 7Документ16 страницE0 UoE Unit 7Patrick GutierrezОценок пока нет
- The JHipster Mini Book 2Документ129 страницThe JHipster Mini Book 2tyulist100% (1)
- Laboratory Manual (CIV 210) Engineering Surveying (2018-19) (For Private Circulation Only)Документ76 страницLaboratory Manual (CIV 210) Engineering Surveying (2018-19) (For Private Circulation Only)gyanendraОценок пока нет
- Nama: Yossi Tiara Pratiwi Kelas: X Mis 1 Mata Pelajaran: Bahasa InggrisДокумент2 страницыNama: Yossi Tiara Pratiwi Kelas: X Mis 1 Mata Pelajaran: Bahasa InggrisOrionj jrОценок пока нет