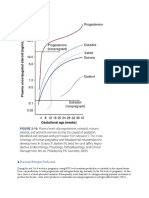
Вам также может понравиться
- Never Split the Difference: Negotiating As If Your Life Depended On ItОт EverandNever Split the Difference: Negotiating As If Your Life Depended On ItРейтинг: 4.5 из 5 звезд4.5/5 (838)
- BPA PCaДокумент12 страницBPA PCaWesley MuhammadОценок пока нет
- AdrosteroneДокумент4 страницыAdrosteroneWesley MuhammadОценок пока нет
- Aromatase and BPAДокумент7 страницAromatase and BPAWesley MuhammadОценок пока нет
- Adams 1Документ5 страницAdams 1Wesley MuhammadОценок пока нет
- ArticleДокумент12 страницArticleWesley MuhammadОценок пока нет
- Drugs 8Документ10 страницDrugs 8Wesley MuhammadОценок пока нет
- Hyperserotonemia in Autism: The Potential Role of 5HT-related Gene VariantsДокумент6 страницHyperserotonemia in Autism: The Potential Role of 5HT-related Gene VariantsWesley MuhammadОценок пока нет
- Bpa 1Документ9 страницBpa 1Wesley MuhammadОценок пока нет
- Black Men Testosterone 14Документ7 страницBlack Men Testosterone 14Wesley MuhammadОценок пока нет
- Black Maculinity 1Документ31 страницаBlack Maculinity 1Wesley Muhammad100% (1)
- Drugs 7Документ6 страницDrugs 7Wesley MuhammadОценок пока нет
- Afrocentricity 1Документ18 страницAfrocentricity 1Wesley MuhammadОценок пока нет
- William HerrmannДокумент68 страницWilliam HerrmannWesley MuhammadОценок пока нет
- 19770523Документ16 страниц19770523Wesley MuhammadОценок пока нет
- 19770523Документ16 страниц19770523Wesley MuhammadОценок пока нет
- Amygdala: Eyes Wide Open: Current Biology Vol 24 No 20 R1000Документ3 страницыAmygdala: Eyes Wide Open: Current Biology Vol 24 No 20 R1000Wesley MuhammadОценок пока нет
- AggressionДокумент16 страницAggressionWesley MuhammadОценок пока нет
- Svayambhu PuranaДокумент9 страницSvayambhu PuranaMin Bahadur shakya88% (8)
- Autism 1Документ13 страницAutism 1Wesley MuhammadОценок пока нет
- South African Archaeological SocietyДокумент15 страницSouth African Archaeological SocietyWesley MuhammadОценок пока нет
- Blond, Tall, With Honey-Colored Eyes Jewish Ownership of Slaves in The Ottoman EmpireДокумент19 страницBlond, Tall, With Honey-Colored Eyes Jewish Ownership of Slaves in The Ottoman EmpireBlack SunОценок пока нет
- Ra IZ Allah Status Quaestionis ReportДокумент30 страницRa IZ Allah Status Quaestionis ReportWesley Muhammad100% (4)
- Skeleton From Iran CaveДокумент13 страницSkeleton From Iran CaveWesley MuhammadОценок пока нет
- Jonathan Smith 1Документ26 страницJonathan Smith 1Wesley MuhammadОценок пока нет
- Buddha GodДокумент3 страницыBuddha GodWesley MuhammadОценок пока нет
- Avrospatt Sacred Origins of The Svayambhucaitya JNRC 13 2009 PDFДокумент58 страницAvrospatt Sacred Origins of The Svayambhucaitya JNRC 13 2009 PDFGB ShresthaОценок пока нет
- Great Buddha 3Документ8 страницGreat Buddha 3Wesley MuhammadОценок пока нет
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeОт EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeРейтинг: 4 из 5 звезд4/5 (5794)
- The Yellow House: A Memoir (2019 National Book Award Winner)От EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Рейтинг: 4 из 5 звезд4/5 (98)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceОт EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceРейтинг: 4 из 5 звезд4/5 (894)
- The Little Book of Hygge: Danish Secrets to Happy LivingОт EverandThe Little Book of Hygge: Danish Secrets to Happy LivingРейтинг: 3.5 из 5 звезд3.5/5 (399)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureОт EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureРейтинг: 4.5 из 5 звезд4.5/5 (474)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryОт EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryРейтинг: 3.5 из 5 звезд3.5/5 (231)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaОт EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaРейтинг: 4.5 из 5 звезд4.5/5 (265)
- The Emperor of All Maladies: A Biography of CancerОт EverandThe Emperor of All Maladies: A Biography of CancerРейтинг: 4.5 из 5 звезд4.5/5 (271)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersОт EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersРейтинг: 4.5 из 5 звезд4.5/5 (344)
- The Unwinding: An Inner History of the New AmericaОт EverandThe Unwinding: An Inner History of the New AmericaРейтинг: 4 из 5 звезд4/5 (45)
- Team of Rivals: The Political Genius of Abraham LincolnОт EverandTeam of Rivals: The Political Genius of Abraham LincolnРейтинг: 4.5 из 5 звезд4.5/5 (234)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyОт EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyРейтинг: 3.5 из 5 звезд3.5/5 (2219)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreОт EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreРейтинг: 4 из 5 звезд4/5 (1090)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)От EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Рейтинг: 4.5 из 5 звезд4.5/5 (119)
- DHEA PrasteronaДокумент125 страницDHEA PrasteronaMiguel Angel Contreras Ramos100% (1)
- Multiple Choice Questions on Androgen Production and EffectsДокумент4 страницыMultiple Choice Questions on Androgen Production and EffectsDrFarah Emad AliОценок пока нет
- Anabolic Steroid Control Act 2004Документ45 страницAnabolic Steroid Control Act 2004Kane SmithОценок пока нет
- Congenital Adrenal HyperplasiaДокумент17 страницCongenital Adrenal HyperplasiaJeff CrocombeОценок пока нет
- Biosynthesis of Steroid Hormones: Dr. Sumreena Mansoor Assistant Prof of BiochemistryДокумент49 страницBiosynthesis of Steroid Hormones: Dr. Sumreena Mansoor Assistant Prof of BiochemistryYumnah BabarОценок пока нет
- Influence of Steroid Hormones On The PeriodontiumДокумент69 страницInfluence of Steroid Hormones On The PeriodontiumSamra NausheenОценок пока нет
- 5450924Документ121 страница5450924Ryan DivishОценок пока нет
- Clinical Evaluation of Purified Shilajit On TestosteroneДокумент6 страницClinical Evaluation of Purified Shilajit On TestosteroneAlejandro MüllerОценок пока нет
- Menopause Symptoms, Causes, and Treatments ExplainedДокумент35 страницMenopause Symptoms, Causes, and Treatments ExplainedCALVO, JUAN BERNARDOОценок пока нет
- Biosintesis Dan Mekanisme Kerja Hormon Repro Wanita PDFДокумент39 страницBiosintesis Dan Mekanisme Kerja Hormon Repro Wanita PDFsallynorcelinaОценок пока нет
- Sports and fitness tips for a healthy lifestyleДокумент24 страницыSports and fitness tips for a healthy lifestyleSahooashuОценок пока нет
- Androgenic Supplementation in Men - Effects of Age Herbal ExtractДокумент154 страницыAndrogenic Supplementation in Men - Effects of Age Herbal ExtractMaria PopaОценок пока нет
- 34 (Tribulus Terrestris) JMPR-11-138 PDFДокумент5 страниц34 (Tribulus Terrestris) JMPR-11-138 PDFmakram_0451Оценок пока нет
- Dehydroepiandrosterone (DHEA) : Hypes and HopesДокумент13 страницDehydroepiandrosterone (DHEA) : Hypes and HopesFernanda PérezОценок пока нет
- Barbell 55 Nutrition Plan by Guru MannДокумент2 страницыBarbell 55 Nutrition Plan by Guru MannVikas TomarОценок пока нет
- A Mans Guide To Testostrone Replacement Therapy PDFДокумент21 страницаA Mans Guide To Testostrone Replacement Therapy PDFdineshmarginalОценок пока нет
- The Steroidogenic PathwayДокумент47 страницThe Steroidogenic Pathwaysusan64Оценок пока нет
- TribullusДокумент6 страницTribullusGREESTYОценок пока нет
- Accelerated Episodic Luteinizing Hormone Release Accompanies Blunted Progesterone Regulation in PCOS-like Female Rhesus Monkeys (Macaca Mulatta) Exposed To Testosterone During Early-to-Mid GestationДокумент14 страницAccelerated Episodic Luteinizing Hormone Release Accompanies Blunted Progesterone Regulation in PCOS-like Female Rhesus Monkeys (Macaca Mulatta) Exposed To Testosterone During Early-to-Mid GestationEricko Ongko JoyoОценок пока нет
- The PDR Family Guide To Nutritional Supplements An Authoritative A To Z Resource On The 100 Most Popular Nutritional Therapies and Nutraceuticals PDДокумент353 страницыThe PDR Family Guide To Nutritional Supplements An Authoritative A To Z Resource On The 100 Most Popular Nutritional Therapies and Nutraceuticals PDtfashourОценок пока нет
- Peripheral Testosterone MetabolismДокумент8 страницPeripheral Testosterone MetabolismAgung SentosaОценок пока нет
- Draft Notice Comments Guidance Doc Supplements Sportspersons 16-03-2018Документ27 страницDraft Notice Comments Guidance Doc Supplements Sportspersons 16-03-2018Kavita YadavОценок пока нет
- Female Reproductive Physiology: Pradip Kumar Das, Joydip Mukherjee, and Dipak BanerjeeДокумент56 страницFemale Reproductive Physiology: Pradip Kumar Das, Joydip Mukherjee, and Dipak BanerjeeVFJulianОценок пока нет
- Sex Hormone Synthesis, Regulation, and Function - McMaster Pathophysiology ReviewДокумент7 страницSex Hormone Synthesis, Regulation, and Function - McMaster Pathophysiology ReviewWahyu W'cas PamungkasОценок пока нет
- Estrogen DR DeanДокумент8 страницEstrogen DR DeanMochammad Halim NОценок пока нет
- NRDP 201529Документ21 страницаNRDP 201529Zurielle LuboaОценок пока нет
- Impact of Extraction Solvents On Steroid Contents Determined in BeefДокумент6 страницImpact of Extraction Solvents On Steroid Contents Determined in BeefMuhammad ZafriОценок пока нет
- The Biochemistry of Hormones ClassificationДокумент45 страницThe Biochemistry of Hormones ClassificationBadini ChanalОценок пока нет
- Upgrade Your TestosteroneДокумент115 страницUpgrade Your TestosteroneKevin100% (3)
- Androgens and Anabolic SteroidsДокумент37 страницAndrogens and Anabolic SteroidsRezy Arina PutriОценок пока нет