Вам также может понравиться
- Ciclo de Krebs: Reacciones bioquímicas para liberar energía en organismos aerobiosДокумент16 страницCiclo de Krebs: Reacciones bioquímicas para liberar energía en organismos aerobiosYuli GarcíaОценок пока нет
- Cadena transporte electrones generación ATPДокумент17 страницCadena transporte electrones generación ATPPaxel LIОценок пока нет
- Evolucion Sistematica y Taxonomia MicrobianaДокумент33 страницыEvolucion Sistematica y Taxonomia Microbianaeliza humpiriОценок пока нет
- Baterias Bioquimicas EnterobacteriasДокумент6 страницBaterias Bioquimicas EnterobacteriasDarinka Nicole TxaipeОценок пока нет
- Urea kit optimizado paraДокумент2 страницыUrea kit optimizado paraFelipe MuñosОценок пока нет
- Metabolismo de Los Carbohidratos, RESUMENДокумент12 страницMetabolismo de Los Carbohidratos, RESUMENpaolaОценок пока нет
- Tinción simple y Gram: identificación de bacteriasДокумент11 страницTinción simple y Gram: identificación de bacteriasliderОценок пока нет
- Acidos GrasosДокумент52 страницыAcidos Grasosmirna victoria reynoso lopezОценок пока нет
- Identificación de CarbohidratosДокумент24 страницыIdentificación de Carbohidratosanon_702017160Оценок пока нет
- Gc3a9nero CorynebacteriumДокумент32 страницыGc3a9nero CorynebacteriumLynda Túllume SeguraОценок пока нет
- Practica de EnzimasДокумент5 страницPractica de EnzimasMayte VenturaОценок пока нет
- Microscopio de Fluorescencia PDFДокумент2 страницыMicroscopio de Fluorescencia PDFJuliana GomezОценок пока нет
- Act12 de BioquimicaДокумент5 страницAct12 de BioquimicaLucy Judith Cortes MartinezОценок пока нет
- Beta oxidación de ácidos grasos insaturados y saturados de cadena parДокумент24 страницыBeta oxidación de ácidos grasos insaturados y saturados de cadena parGise HinostrozaОценок пока нет
- RESULTADOS Cocos Gram PositivosДокумент7 страницRESULTADOS Cocos Gram PositivosJaime Diaz PedrozoОценок пока нет
- Nutrición bacteriana y metabolismoДокумент58 страницNutrición bacteriana y metabolismoEsteban Alejandro Bertarelli OteroОценок пока нет
- Metabolismo Del Glucogeno (Presentación)Документ27 страницMetabolismo Del Glucogeno (Presentación)Guadalupe RiveraОценок пока нет
- AGAR HIERRO-LISINA DIFERENCIA ENTEROBACTERIASДокумент17 страницAGAR HIERRO-LISINA DIFERENCIA ENTEROBACTERIASOscar Zamora100% (1)
- La GlucólisisДокумент27 страницLa Glucólisisrock2312Оценок пока нет
- Problemas de genética mendelianaДокумент3 страницыProblemas de genética mendelianaNancy Lopez cervantesОценок пока нет
- Bioquimica 1. Analisis Cuantitativo de EnzimasДокумент12 страницBioquimica 1. Analisis Cuantitativo de EnzimasangelaОценок пока нет
- Glucolisis Anaerobia (Preguntas) Lab.Документ4 страницыGlucolisis Anaerobia (Preguntas) Lab.Maleja SuarezОценок пока нет
- Identificación de Sustancias Orgánicas y Determinación de EstructurasДокумент9 страницIdentificación de Sustancias Orgánicas y Determinación de EstructurasVanesa VanessОценок пока нет
- Fundamento y Principios Del Metodo de ElisaДокумент12 страницFundamento y Principios Del Metodo de ElisaVivi SaldiviaОценок пока нет
- Cuestionario Carboxilicos y Ruta de Las PentosasДокумент2 страницыCuestionario Carboxilicos y Ruta de Las PentosasAlfonso Lopez PeñafordОценок пока нет
- Ejercicio N°12 - Bioquímica - OCWДокумент5 страницEjercicio N°12 - Bioquímica - OCWYan HernándezОценок пока нет
- Metabolismo Del NitrogenoДокумент36 страницMetabolismo Del NitrogenoGeovanni moralesОценок пока нет
- Alimentos FuncionalesДокумент41 страницаAlimentos FuncionalesFriedberd MarioОценок пока нет
- Cuestionario de Glucidos y MetabolismoДокумент5 страницCuestionario de Glucidos y MetabolismoIAGO SAMUEL DE SOUZA REISОценок пока нет
- Fosforilacion OxidativaДокумент16 страницFosforilacion OxidativaFlorencia BarsiОценок пока нет
- Proteinas de CoagulacionДокумент31 страницаProteinas de CoagulacionHematoped 20 novОценок пока нет
- Fosforilacion Oxidativa GatoДокумент12 страницFosforilacion Oxidativa GatoCarmelita Hcs RmrezОценок пока нет
- Estrategia Catalítica de La LisozimaДокумент17 страницEstrategia Catalítica de La LisozimaLudwig Omar Julca Salazar100% (4)
- YURIANIS BANDA - Taller+de+Química+Medicinal SEMANA 3Документ6 страницYURIANIS BANDA - Taller+de+Química+Medicinal SEMANA 3anon_287131257Оценок пока нет
- Estracción de CaseinaДокумент4 страницыEstracción de Caseinamaria stella ortega suarezОценок пока нет
- Sintesis de PirimidinaДокумент4 страницыSintesis de PirimidinaBrendaBelloОценок пока нет
- LipidPeroxidación3PhaseProcessДокумент3 страницыLipidPeroxidación3PhaseProcessEduardo LobitoОценок пока нет
- Esquemas de MetabolismoДокумент5 страницEsquemas de MetabolismoSandra CardosoОценок пока нет
- Sesion 08. - Guia de Ejercicios N°02Документ45 страницSesion 08. - Guia de Ejercicios N°02riesgos33100% (1)
- Guia 5 - Permeabilidad Celular - ERITROCITOS Y ELODEAДокумент8 страницGuia 5 - Permeabilidad Celular - ERITROCITOS Y ELODEANIKOLE SANABRIA RODRIGUEZОценок пока нет
- Factores Quimiotacticos EndogenosДокумент9 страницFactores Quimiotacticos EndogenosDIEGO ALEJANDRO GARCIA RINCONОценок пока нет
- RIFAMICINAДокумент10 страницRIFAMICINAAngel Daniel DominguezОценок пока нет
- Reactivo de BenedictДокумент2 страницыReactivo de BenedictDaniel'sAcosta60% (5)
- Práctica 7. Digestión de ProteínasДокумент5 страницPráctica 7. Digestión de ProteínasSelvis Rivera CerezoОценок пока нет
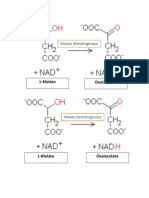
- La Enzima Malato DeshidrogenasaДокумент2 страницыLa Enzima Malato DeshidrogenasaKaren Torres JuárezОценок пока нет
- Guiatecnologiafarmaceuticaii2013 130306193107 Phpapp01 PDFДокумент104 страницыGuiatecnologiafarmaceuticaii2013 130306193107 Phpapp01 PDFGUILLERMOОценок пока нет
- Metabolismo Celular 2017Документ44 страницыMetabolismo Celular 2017juan pa100% (1)
- Activación de Los Ácidos GrasosДокумент2 страницыActivación de Los Ácidos GrasosJaviera Gotuzzo CarrascoОценок пока нет
- Cuestionario de Fisiología y Metabolismo MicrobianoДокумент2 страницыCuestionario de Fisiología y Metabolismo MicrobianoLaura JinenezОценок пока нет
- Cromatografía de Filtración en Gel 1Документ6 страницCromatografía de Filtración en Gel 1Jenny Luz GomezОценок пока нет
- Expresión y regulación génica en procariotasДокумент36 страницExpresión y regulación génica en procariotasAbner AlvaradoОценок пока нет
- Protocolo Tincion CFU-F GiemsaДокумент4 страницыProtocolo Tincion CFU-F GiemsaAtripoloneОценок пока нет
- Transporte de Cargas en Sistemas BiologicosДокумент2 страницыTransporte de Cargas en Sistemas Biologicosyoemis100% (1)
- Cadena Respiratoria Fosforilacion OxidatДокумент95 страницCadena Respiratoria Fosforilacion OxidatMiguel Soni CasillasОценок пока нет
- Lehninger, A. Cap06 EnzimasДокумент48 страницLehninger, A. Cap06 EnzimasTanilvalОценок пока нет
- Farmacognosia GB - 15Документ12 страницFarmacognosia GB - 15Liendo Polanco GustavoОценок пока нет
- Alcohol EsДокумент15 страницAlcohol EsBastian Muñoz RobledoОценок пока нет
- Ciclo de Krebs 2023Документ39 страницCiclo de Krebs 2023Jennifer González100% (1)
- Tema 11. Metabolismo Del Piruvato PDFДокумент32 страницыTema 11. Metabolismo Del Piruvato PDFALBA CONCHUELA ARENASОценок пока нет
- Lomm080123hdfprra0 PDFДокумент1 страницаLomm080123hdfprra0 PDFJm AlmarazОценок пока нет
- Presentacion Empleo Covid19 CDMX Styfe PDFДокумент12 страницPresentacion Empleo Covid19 CDMX Styfe PDFJm AlmarazОценок пока нет
- Icrc 003 0880Документ170 страницIcrc 003 0880MotorolaBooksОценок пока нет
- DiplomaДокумент1 страницаDiplomaJm AlmarazОценок пока нет
- Acordeon Extra DinamicaДокумент7 страницAcordeon Extra DinamicaJm AlmarazОценок пока нет
- Icrc 003 0880Документ170 страницIcrc 003 0880MotorolaBooksОценок пока нет
- Ricos HogДокумент4 страницыRicos HogJm AlmarazОценок пока нет
- NSMaterial DidacticoДокумент148 страницNSMaterial DidacticoJm AlmarazОценок пока нет
- NSMaterial DidacticoДокумент148 страницNSMaterial DidacticoJm AlmarazОценок пока нет
- NSMaterial DidacticoДокумент148 страницNSMaterial DidacticoJm AlmarazОценок пока нет
- FormatosДокумент6 страницFormatosJm AlmarazОценок пока нет
- TarjetaДокумент1 страницаTarjetaJm AlmarazОценок пока нет
- Mini InvestigaciónДокумент13 страницMini InvestigaciónJm AlmarazОценок пока нет
- Actualiza InfoДокумент3 страницыActualiza InfoJm AlmarazОценок пока нет
- Carta Colegio de México PDFДокумент1 страницаCarta Colegio de México PDFJm AlmarazОценок пока нет
- Meade Propone Que Sean Deducibles Colegiaturas de Nivel SuperiorДокумент1 страницаMeade Propone Que Sean Deducibles Colegiaturas de Nivel SuperiorJm AlmarazОценок пока нет
- DP Eje1-Gas NaturalДокумент3 страницыDP Eje1-Gas NaturalJm AlmarazОценок пока нет
- Gilberto Enrique Díaz CastroДокумент1 страницаGilberto Enrique Díaz CastroJm AlmarazОценок пока нет
- ConferenciaДокумент4 страницыConferenciaJm AlmarazОценок пока нет
- Diploma - Docx - Documentos de GoogleДокумент1 страницаDiploma - Docx - Documentos de GoogleJm AlmarazОценок пока нет
- ConferenciaДокумент4 страницыConferenciaJm AlmarazОценок пока нет
- Reposición 3Документ1 страницаReposición 3Jm AlmarazОценок пока нет
- Contrib Uc I OnesДокумент8 страницContrib Uc I OnesJm AlmarazОценок пока нет
- PlaneacionДокумент18 страницPlaneacionJm AlmarazОценок пока нет
- Des Arrollo Equipo S 22 A BRДокумент1 страницаDes Arrollo Equipo S 22 A BRJm AlmarazОценок пока нет
- Lonas PDFДокумент1 страницаLonas PDFJm AlmarazОценок пока нет
- Proceso Elaboracion Enlatado Atun AguaДокумент50 страницProceso Elaboracion Enlatado Atun Aguageorgeth21100% (1)
- Olimpiada Del Saber III Periodo NovenoДокумент2 страницыOlimpiada Del Saber III Periodo NovenoKatherine Cedeño PuentesОценок пока нет
- FamiliaДокумент12 страницFamiliaEliudAbrahamОценок пока нет
- APORTES DE LA BIOLOGIA A LA PSICOLOGIA 3.jpegДокумент1 страницаAPORTES DE LA BIOLOGIA A LA PSICOLOGIA 3.jpeggeraldint cetinaОценок пока нет
- Horario para Los Padres2023Документ1 страницаHorario para Los Padres2023mario toledoОценок пока нет
- Santiago (Ica) - Metodologia de La InvestigacionДокумент42 страницыSantiago (Ica) - Metodologia de La InvestigacionYhonny Arce Romero0% (1)
- Línea Del Tiempo Psicología GeneralДокумент11 страницLínea Del Tiempo Psicología GeneralAngelica Ferras50% (2)
- Celulas AnimalesДокумент2 страницыCelulas AnimalesVania ReynosoОценок пока нет
- Reglamento Construcción San Matias 2013-11-01 FINALДокумент28 страницReglamento Construcción San Matias 2013-11-01 FINALAlfredo OrfilaОценок пока нет
- Redes de TriangulaciónДокумент10 страницRedes de Triangulaciónlenin smith dominguez dominguezОценок пока нет
- Decreto 606 de 2001Документ38 страницDecreto 606 de 2001Oscar Daniel Valencia GonzalezОценок пока нет
- Memoria Descriptiva Del Plano Perimetrico de PoligonosДокумент3 страницыMemoria Descriptiva Del Plano Perimetrico de PoligonosJhonny KenОценок пока нет
- Determinacion de Rumbo y Azimut de Una Poligonal Abierta2Документ15 страницDeterminacion de Rumbo y Azimut de Una Poligonal Abierta2Juan Villarreal SalomeОценок пока нет
- El Pleistoceno y HolosenoДокумент1 страницаEl Pleistoceno y HolosenoAndersson Julca Vasquez100% (1)
- Lensometria Eh 2021Документ84 страницыLensometria Eh 2021Daymler ivan alcazar landa100% (3)
- Metabolitos Primarios y Secundarios - Clase 3Документ27 страницMetabolitos Primarios y Secundarios - Clase 3Maritza MPОценок пока нет
- Determinacion Del NorteДокумент24 страницыDeterminacion Del NorteJose Almanza100% (2)
- Arnold Krumm Heller - Astrologia PDFДокумент36 страницArnold Krumm Heller - Astrologia PDFMiguel Ángel Blanco VegaОценок пока нет
- Lischetti 2Документ2 страницыLischetti 2Daniela Gauna LujanОценок пока нет
- SNB Programa y Cronograma de Sustentabilidad Del Medio AmbienteДокумент13 страницSNB Programa y Cronograma de Sustentabilidad Del Medio AmbienteCris Riffo CerdaОценок пока нет
- Evaluación de Fármacos y Medicamentos IДокумент2 страницыEvaluación de Fármacos y Medicamentos IPao MoralesОценок пока нет
- El Rio AmazonasДокумент9 страницEl Rio AmazonasAlicia Mamani YagunoОценок пока нет
- Cuatro Libros de La Naturaleza y Virtudes Medicinales de Las Plantas y Animales de La Nueva EspañaДокумент390 страницCuatro Libros de La Naturaleza y Virtudes Medicinales de Las Plantas y Animales de La Nueva EspañaSalvador Alcantar0% (1)
- 3 Basico - Cs. Naturales - Norma - EstudianteДокумент122 страницы3 Basico - Cs. Naturales - Norma - EstudianteCarla Bugueño DíazОценок пока нет
- Hds-Sulfato de Amonio (Honeywell)Документ6 страницHds-Sulfato de Amonio (Honeywell)Eloy Perez ValeraОценок пока нет
- A Hombro de GigantesДокумент8 страницA Hombro de Gigantesabner_chingaОценок пока нет
- PNLДокумент16 страницPNLLuna Tamara KantorОценок пока нет
- Practico #3 Fuerza y EstáticaДокумент2 страницыPractico #3 Fuerza y EstáticaErck PattonОценок пока нет
- Hidruros Miguel Ángel Lorenzo Fossati 3º A PDFДокумент2 страницыHidruros Miguel Ángel Lorenzo Fossati 3º A PDFMaribel FossatiОценок пока нет
- Cuestionario Ley de ArchivosДокумент10 страницCuestionario Ley de ArchivosfrancoОценок пока нет