Вам также может понравиться
- Cytogenetics Midterm2Документ4 страницыCytogenetics Midterm2Nikoh Anthony EwayanОценок пока нет
- Duplications (Dup) : Direct InvertedДокумент22 страницыDuplications (Dup) : Direct Invertedmaham javaidОценок пока нет
- Human Chromosome NomenclatureДокумент35 страницHuman Chromosome NomenclatureBabak Nami91% (22)
- Chapter4 CytogeneticsДокумент70 страницChapter4 CytogeneticsAIMAN MUHAIMMIN HASNANОценок пока нет
- Human Genetics 2Документ4 страницыHuman Genetics 2ll2997271Оценок пока нет
- Chapter 3Документ7 страницChapter 3candra mahardikaОценок пока нет
- Sex Chromosome Aberrations and Transsexualism: VOL. 79, NO. 3, MARCH 2003Документ2 страницыSex Chromosome Aberrations and Transsexualism: VOL. 79, NO. 3, MARCH 2003Fvg Fvg FvgОценок пока нет
- Cytogenetic Practice I: No. 435-Course: HeredityДокумент21 страницаCytogenetic Practice I: No. 435-Course: HeredityEdgardo Aguilar HernandezОценок пока нет
- Chromosomal Theory of InheritanceДокумент52 страницыChromosomal Theory of Inheritanceamairahdee304Оценок пока нет
- 2 Chromosomal AbnormalitiesДокумент26 страниц2 Chromosomal Abnormalitiesmuhammad zakriaОценок пока нет
- KaryotypingДокумент21 страницаKaryotypingSarthakОценок пока нет
- KaryotypeДокумент15 страницKaryotypeMustafa Khandgawi0% (1)
- HeredityДокумент39 страницHereditySamantha CunananОценок пока нет
- (Lect 1) Karyotyping - Chromosomal AbnormalitiesДокумент31 страница(Lect 1) Karyotyping - Chromosomal Abnormalitiesshathaehab86Оценок пока нет
- Chromosome Translocations and CancerДокумент5 страницChromosome Translocations and CancervictoryОценок пока нет
- Sitogenetika&variasi GenetikДокумент147 страницSitogenetika&variasi GenetikDiniОценок пока нет
- Human CytogeneticДокумент35 страницHuman CytogeneticanibandasoОценок пока нет
- M3EA1 Karyotyping Group WorkДокумент11 страницM3EA1 Karyotyping Group WorklivОценок пока нет
- 2 GeneticsДокумент23 страницы2 GeneticszahuuОценок пока нет
- ChromosomesДокумент4 страницыChromosomesعامر جدونОценок пока нет
- Aneuploidy GeneticsДокумент7 страницAneuploidy Geneticslinh myОценок пока нет
- CYTOGENETICS Lesson 2 Intro To CytologyДокумент6 страницCYTOGENETICS Lesson 2 Intro To CytologyC AccountОценок пока нет
- Chromosomal AbnormalitiesДокумент42 страницыChromosomal AbnormalitiesAnkit VermaОценок пока нет
- 11 Human Cy To Genetics TalkДокумент11 страниц11 Human Cy To Genetics Talksami95Оценок пока нет
- KaryotypeДокумент9 страницKaryotypeAppas SahaОценок пока нет
- Structural Abnormalities of ChromosomesДокумент31 страницаStructural Abnormalities of ChromosomesAimst GirishprabhuОценок пока нет
- Cytogenetics FLIPPED CLASS 2021-02 3Документ19 страницCytogenetics FLIPPED CLASS 2021-02 3Diran TerzyanОценок пока нет
- Cytogenetics Note PDFДокумент14 страницCytogenetics Note PDFMerjema Bahtanović100% (1)
- Cytogenetics in ReproductionДокумент23 страницыCytogenetics in ReproductionmagdecitaОценок пока нет
- Introduction To CytogeneticsДокумент25 страницIntroduction To CytogeneticsFelix John Paul Barqueros100% (1)
- WEEK 11 & 12 (Part 1) (CYTO)Документ2 страницыWEEK 11 & 12 (Part 1) (CYTO)Hershei Vonne BaccayОценок пока нет
- 2014 Genetics Lecture M Kamal PDFДокумент93 страницы2014 Genetics Lecture M Kamal PDFapi-255205775Оценок пока нет
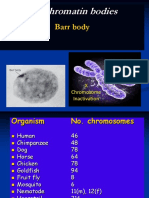
- Sex Chromatin Bodies: Barr BodyДокумент45 страницSex Chromatin Bodies: Barr BodyKhaleelMetlakОценок пока нет
- ChromosomesДокумент9 страницChromosomesmm3370350Оценок пока нет
- Karyotyping ActivityДокумент4 страницыKaryotyping ActivityYSOBELLATHERESE BILOLOОценок пока нет
- CytogeneticsДокумент5 страницCytogeneticsAndriaОценок пока нет
- Centromere. Depending On The Centromere Location, The ChromosomesДокумент5 страницCentromere. Depending On The Centromere Location, The ChromosomesNargesОценок пока нет
- Bio511 c2Документ51 страницаBio511 c2Athirah JeffryОценок пока нет
- Cytogenetics: Chromosome Banding & Barr Body DR Charu Khosla, 24/4/20, 10.45-11.45 AmДокумент90 страницCytogenetics: Chromosome Banding & Barr Body DR Charu Khosla, 24/4/20, 10.45-11.45 Amsonal aranha100% (2)
- Genetic Linkage: Presented By:ayush Jain (Alm 3008) University of Agricultural Science, BangloreДокумент32 страницыGenetic Linkage: Presented By:ayush Jain (Alm 3008) University of Agricultural Science, BangloreTeflon SlimОценок пока нет
- Chapt 24Документ0 страницChapt 24SilaxОценок пока нет
- 8.2 GMppt-chapter5-2-2020 (Ok and Important)Документ41 страница8.2 GMppt-chapter5-2-2020 (Ok and Important)Kw ChanОценок пока нет
- Chromosomal MutationДокумент7 страницChromosomal Mutationchastine daneОценок пока нет
- 7 Linkage MappingДокумент86 страниц7 Linkage MappingEmi JiHyeon KimОценок пока нет
- Genetika 2Документ21 страницаGenetika 2Sreckovic BenjoОценок пока нет
- S.Y.B.Sc Semester III Botany Paper II Unit II:CytogeneticsДокумент95 страницS.Y.B.Sc Semester III Botany Paper II Unit II:CytogeneticsQueen1982 K100% (1)
- Genetics Minimals FinalДокумент5 страницGenetics Minimals FinalМөнхгэрэл ГанбатОценок пока нет
- KaryotypeДокумент11 страницKaryotypeafzalОценок пока нет
- 2.linkage Crossing Over and RecombinationДокумент101 страница2.linkage Crossing Over and RecombinationChandrachur GhoshОценок пока нет
- Choose The Correct AnswerДокумент8 страницChoose The Correct AnswerBLlahniesss Love KenОценок пока нет
- ChroMutations F05Документ24 страницыChroMutations F05Homes EreñoОценок пока нет
- Diagnosis of Four Chromosome Abnormalities of Unknown Origin Chromosome Microdissection and Subsequent Reverse and Forward PaintingДокумент4 страницыDiagnosis of Four Chromosome Abnormalities of Unknown Origin Chromosome Microdissection and Subsequent Reverse and Forward PaintingJosé Abraham Amaya DuarteОценок пока нет
- Genetic Diversity in The UV Sex Chromosomes of TheДокумент18 страницGenetic Diversity in The UV Sex Chromosomes of TheCarlos Lesmes DíazОценок пока нет
- 4 Genetics Lecture - Karyotyping of Normal Male, Female and Different Chromosomal Anomalies.Документ43 страницы4 Genetics Lecture - Karyotyping of Normal Male, Female and Different Chromosomal Anomalies.AMIRA HELAYELОценок пока нет
- Chapter 2 GeneticsДокумент45 страницChapter 2 Geneticskristinenidoy.mdОценок пока нет
- Soma ( Body)Документ4 страницыSoma ( Body)Joe AjaОценок пока нет
- Chromosome StructureДокумент11 страницChromosome StructureAshley Nicole DelizoОценок пока нет
- 2 ChromosomeДокумент37 страниц2 ChromosomeSarveashini SegarОценок пока нет
- Genome Editing-Medical-PowerPoint-TemplatesДокумент23 страницыGenome Editing-Medical-PowerPoint-TemplatesLolita SyahnazОценок пока нет
- Flock MenuДокумент5 страницFlock MenuWilson TayОценок пока нет
- ISKCON Desire Tree - Brahma Vimohana LeelaДокумент34 страницыISKCON Desire Tree - Brahma Vimohana LeelaISKCON desire treeОценок пока нет
- Ableism - What It Is and Why It Matters To EveryoneДокумент28 страницAbleism - What It Is and Why It Matters To Everyonellemma admasОценок пока нет
- EVK203/EVK213/EVK223/EVK233/EVK253: Digital Thermostats For Ventilated Refrigerating UnitsДокумент2 страницыEVK203/EVK213/EVK223/EVK233/EVK253: Digital Thermostats For Ventilated Refrigerating UnitsMihai BordeianuОценок пока нет
- VW Golf 8 Variant WD EngДокумент664 страницыVW Golf 8 Variant WD EngLakhdar BouchenakОценок пока нет
- Biological Activity of Bone Morphogenetic ProteinsДокумент4 страницыBiological Activity of Bone Morphogenetic Proteinsvanessa_werbickyОценок пока нет
- Testing For Cations Flow ChartДокумент2 страницыTesting For Cations Flow Chartapi-252561013Оценок пока нет
- Final Workshop Report On Value Addition and AgroprocessingДокумент31 страницаFinal Workshop Report On Value Addition and AgroprocessingBett K. BernardОценок пока нет
- Enzymes in Grain ProcessingДокумент1 страницаEnzymes in Grain ProcessingAttila-Levente FogarasiОценок пока нет
- EcodesДокумент2 страницыEcodesValentin IonutОценок пока нет
- Scala: Service InstructionsДокумент16 страницScala: Service Instructionsmario_turbinadoОценок пока нет
- Glycerin As A Renewable Feedstock For EpichlorohydДокумент6 страницGlycerin As A Renewable Feedstock For EpichlorohydMuhammad RidhauddinОценок пока нет
- D8045-16 Standard Test Method For Acid Number of Crude Oils and Petroleum Products by CatalyticДокумент6 страницD8045-16 Standard Test Method For Acid Number of Crude Oils and Petroleum Products by CatalytichishamОценок пока нет
- Material SelfДокумент9 страницMaterial Selfic perlasОценок пока нет
- NWO Plans Exposed by Insider in 1969Документ36 страницNWO Plans Exposed by Insider in 1969Stig Dragholm100% (3)
- Eastman Methyl N-Amyl Ketone (MAK) and Eastman Methyl Isoamyl Ketone (MIAK)Документ4 страницыEastman Methyl N-Amyl Ketone (MAK) and Eastman Methyl Isoamyl Ketone (MIAK)Chemtools Chemtools100% (1)
- Full Download Book Microbiome Immunity Digestive Health and Nutrition Epidemiology Pathophysiology Prevention and Treatment PDFДокумент41 страницаFull Download Book Microbiome Immunity Digestive Health and Nutrition Epidemiology Pathophysiology Prevention and Treatment PDFmildred.walker324100% (15)
- 10 Major Sulfuric Acid Industrial Applications - WorldOfChemicalsДокумент9 страниц10 Major Sulfuric Acid Industrial Applications - WorldOfChemicalsFarhad MalikОценок пока нет
- AcuityPDR BrochureДокумент3 страницыAcuityPDR BrochureJulian HutabaratОценок пока нет
- Weigh Bridge Miscellaneous Items DetailsДокумент1 страницаWeigh Bridge Miscellaneous Items DetailsChandan RayОценок пока нет
- I. External Analysis A. General Environment A. Economic DevelopmentsДокумент17 страницI. External Analysis A. General Environment A. Economic DevelopmentsAndrea TaganginОценок пока нет
- Preparation of Stick Type Solid Glue As Paper AdheДокумент9 страницPreparation of Stick Type Solid Glue As Paper AdheAhmad AlShahrourОценок пока нет
- Pathological Anatomy IntroДокумент27 страницPathological Anatomy IntroJoiya KhanОценок пока нет
- InvoiceДокумент1 страницаInvoicesunil sharmaОценок пока нет
- Savage Inequalities Reading ReflectionДокумент2 страницыSavage Inequalities Reading Reflectionapi-367127133Оценок пока нет
- DWATCHДокумент92 страницыDWATCHFawaz SayedОценок пока нет
- Parenting Styles and Social Interaction of Senior Secondary School Students in Imo State, NigeriaДокумент10 страницParenting Styles and Social Interaction of Senior Secondary School Students in Imo State, NigeriaInternational Educational Applied Scientific Research Journal (IEASRJ)Оценок пока нет
- Outback MenuДокумент2 страницыOutback MenuzeeОценок пока нет
- 493M GHC MG Mep WS L3 205 1B 00Документ1 страница493M GHC MG Mep WS L3 205 1B 00shehanОценок пока нет
- Chapter 4 - Medical Aspects of Food SafetyДокумент17 страницChapter 4 - Medical Aspects of Food SafetyasushkОценок пока нет