Вам также может понравиться
- Clase 7 Metabolismo Del Grupo HemoДокумент17 страницClase 7 Metabolismo Del Grupo HemoAguss FernandezОценок пока нет
- Vitaminas, Coenzimas y Derivados de Las VitaminasДокумент12 страницVitaminas, Coenzimas y Derivados de Las VitaminasAntonyОценок пока нет
- Bilirrubina BioquimicaДокумент10 страницBilirrubina BioquimicaJezabel Huaman100% (1)
- HemoglobinaДокумент19 страницHemoglobinaNicolás Ferro PeñuelaОценок пока нет
- Ciclo de La Urea, BioquimicaДокумент5 страницCiclo de La Urea, Bioquimicamaytee19Оценок пока нет
- Resumen de Compuestos NitrogenadosДокумент16 страницResumen de Compuestos NitrogenadosCap Winter 720Оценок пока нет
- Perfil HepáticoДокумент7 страницPerfil HepáticoStephanyChavezFeriaОценок пока нет
- Biosíntesis de La HemoglobinaДокумент10 страницBiosíntesis de La HemoglobinaRosa Pilar Mamani HilasacaОценок пока нет
- Anemia MegaloblasticaДокумент17 страницAnemia MegaloblasticaHarDmando LauraОценок пока нет
- Determinación de Transaminasas en SangreДокумент9 страницDeterminación de Transaminasas en SangreKarime Gonzales RamírezОценок пока нет
- Ciclo de La UreaДокумент65 страницCiclo de La UreaAlexandraMedinaCastilloОценок пока нет
- Practica 2 BilirrubinaДокумент10 страницPractica 2 BilirrubinaJoanie StewartОценок пока нет
- Proteínas TotalesДокумент7 страницProteínas TotalesAndres LlontopОценок пока нет
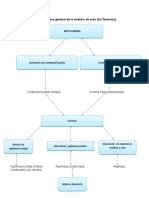
- Esquema General de La Síntesis de UreaДокумент1 страницаEsquema General de La Síntesis de UreaKhaleb Bien AimeОценок пока нет
- Compuestos Nitrogenados No ProteicosДокумент2 страницыCompuestos Nitrogenados No ProteicosPrinci P Real50% (2)
- Determinacion de Proteinas Totales Fraccionadas en SangreДокумент2 страницыDeterminacion de Proteinas Totales Fraccionadas en Sangrejornada fisiologia UNFVОценок пока нет
- Enzimas de Intéres ClínicoДокумент54 страницыEnzimas de Intéres ClínicoMario Gonzalez Olguin100% (2)
- Hemoglobina PDFДокумент9 страницHemoglobina PDFJavi R. HatemОценок пока нет
- Clase Grabada de UreaДокумент18 страницClase Grabada de UreaJuan Diego Leon AguilarОценок пока нет
- Discusion de Grupo N 3Документ14 страницDiscusion de Grupo N 3RubenPerez100% (4)
- Determinaciòn Transaminasa y CreatininaДокумент11 страницDeterminaciòn Transaminasa y CreatininaKarim Merino ObleaОценок пока нет
- Creatinfosfoquinasa MONOGRAFÍAДокумент13 страницCreatinfosfoquinasa MONOGRAFÍAdiego del valleОценок пока нет
- Enfermedad de Orina A Jarabe de ArceДокумент15 страницEnfermedad de Orina A Jarabe de ArceDiana Carolina EscobarОценок пока нет
- Proteinas PlasmaticasДокумент8 страницProteinas PlasmaticasJenniffer Andrea Gordon EstrellaОценок пока нет
- 11º Enzimas Cardíacas y MuscularesДокумент29 страниц11º Enzimas Cardíacas y MuscularesNelidaMamaniОценок пока нет
- Perfil CardiacoДокумент11 страницPerfil CardiacoVero Vero-veroОценок пока нет
- Aspartato AminotransferasaДокумент6 страницAspartato AminotransferasaQarla Yambe ArmentaОценок пока нет
- Desarrollo de ObjetivosДокумент9 страницDesarrollo de ObjetivosMiguel AlvaradoОценок пока нет
- FOSFATASASДокумент13 страницFOSFATASASBîłłý Řôğěř100% (1)
- Fosfatasas AlcalinasДокумент3 страницыFosfatasas AlcalinassofiaОценок пока нет
- Pruebas Bioquimicas PDFДокумент3 страницыPruebas Bioquimicas PDFNatiОценок пока нет
- Metabolismo de Hemoglobina (DV)Документ9 страницMetabolismo de Hemoglobina (DV)Dary Deivy Varela AracheОценок пока нет
- RIBOZIMASДокумент7 страницRIBOZIMASDaniela OlguinОценок пока нет
- Caso Clinico Anemia HemoliticaДокумент10 страницCaso Clinico Anemia HemoliticaIngrid Estefany BamacaОценок пока нет
- Cuestionario de BioquimicaДокумент8 страницCuestionario de BioquimicaBel OcОценок пока нет
- Sintesis de PurinasДокумент5 страницSintesis de PurinasNavaОценок пока нет
- Tecnica de Albumian en SueroДокумент2 страницыTecnica de Albumian en SueroQUIMICO CLINICO WILLIANS SANCHEZ100% (1)
- Metabolismo de LipoproteínasДокумент24 страницыMetabolismo de LipoproteínasPalmira SobalvarroОценок пока нет
- Atlas UrineДокумент2 страницыAtlas UrineScribdTranslationsОценок пока нет
- Aminoacidos No Esenciales (Expo)Документ16 страницAminoacidos No Esenciales (Expo)AlbaOsorioОценок пока нет
- 17 CetosteroidesДокумент2 страницы17 CetosteroidessorangelОценок пока нет
- 24) Ciclo de La UreaДокумент7 страниц24) Ciclo de La UreaHugo Esteban LopezОценок пока нет
- Sintesis de Glicolipidos y Su Función en El OrganismoДокумент9 страницSintesis de Glicolipidos y Su Función en El OrganismoCarlos Dorantes100% (4)
- PRACTICA 9 Proteasa 2014Документ4 страницыPRACTICA 9 Proteasa 2014Ricardo PinillosОценок пока нет
- Metabolismo de La BilirubinaДокумент4 страницыMetabolismo de La BilirubinaNelson Thomas CastroОценок пока нет
- Ciclo de La UreaДокумент10 страницCiclo de La UreaJesus RamirezОценок пока нет
- Metabolismo Del Ácido ÚricoДокумент2 страницыMetabolismo Del Ácido ÚricoBettsy RodríguezОценок пока нет
- Preguntas de Bioquimica AmbientalДокумент3 страницыPreguntas de Bioquimica AmbientalJoe ȺrielОценок пока нет
- Fosfatasa AlcalinaДокумент10 страницFosfatasa AlcalinaluzbnОценок пока нет
- Cinetica de La HemoglobinaДокумент12 страницCinetica de La HemoglobinaCristian SarmientoОценок пока нет
- Metabolismo de Hemoglobina y BilirrubinaДокумент10 страницMetabolismo de Hemoglobina y BilirrubinaJohann FulgencioОценок пока нет
- Fosfatasa 1 y 2Документ14 страницFosfatasa 1 y 2carlosОценок пока нет
- Práctica 13Документ4 страницыPráctica 13Alondra FortunatОценок пока нет
- 7 - Perfil Hepático.....Документ16 страниц7 - Perfil Hepático.....Sergio Diaz CarrascoОценок пока нет
- Morfología Del Eritron Grupo #4Документ37 страницMorfología Del Eritron Grupo #4Michelle Mejia FamiliaОценок пока нет
- AsparaginaДокумент3 страницыAsparaginaMaribel PerezОценок пока нет
- Banco de Preguntas BIOQUIMICAДокумент5 страницBanco de Preguntas BIOQUIMICAThe Master PrintОценок пока нет
- Estudios de Caso Medicina 2019 I - Prof. LeyvaДокумент8 страницEstudios de Caso Medicina 2019 I - Prof. LeyvahenryОценок пока нет
- AminoácidosДокумент9 страницAminoácidosDaniela MoraОценок пока нет
- Apuntes BQДокумент6 страницApuntes BQJulissa Naomi Troya ToalaОценок пока нет
- Cor PulmonaeДокумент17 страницCor PulmonaeJhusleidy España ArroyoОценок пока нет
- RabdomiolisisДокумент8 страницRabdomiolisisJhusleidy España ArroyoОценок пока нет
- ElectrocardiogramaДокумент46 страницElectrocardiogramaJhusleidy España ArroyoОценок пока нет
- CAPÍTULO II - Anormalidades de La PercepcionДокумент10 страницCAPÍTULO II - Anormalidades de La PercepcionJhusleidy España ArroyoОценок пока нет
- Enfermedades Del Almacenamiento de GlucógenoДокумент9 страницEnfermedades Del Almacenamiento de GlucógenoJhusleidy España ArroyoОценок пока нет
- Geriatria y Evaluacion IntegralДокумент43 страницыGeriatria y Evaluacion IntegralJhusleidy España Arroyo100% (1)
- Enfermedades Por D. GlucogenoДокумент19 страницEnfermedades Por D. GlucogenoJhusleidy España ArroyoОценок пока нет
- METABOLISMO DE LOS HC - DiapositivasДокумент25 страницMETABOLISMO DE LOS HC - DiapositivasJhusleidy España ArroyoОценок пока нет
- Cuerpos de Inclusion ViralДокумент1 страницаCuerpos de Inclusion ViralJhusleidy España ArroyoОценок пока нет
- Insuficiencia Respiratoria GuytonДокумент14 страницInsuficiencia Respiratoria GuytonJhusleidy España Arroyo100% (1)
- Interacción de Los Filamentos de Activa y MiosinaДокумент5 страницInteracción de Los Filamentos de Activa y MiosinaJhusleidy España ArroyoОценок пока нет
- Interaccion de Los Filamentos de Actina (Diapositivas)Документ12 страницInteraccion de Los Filamentos de Actina (Diapositivas)Jhusleidy España ArroyoОценок пока нет
- Copywriting - Guía Definitiva para Escribir Textos Que VendenДокумент1 страницаCopywriting - Guía Definitiva para Escribir Textos Que Vendendavid cisnerosОценок пока нет
- Aztecas y NayasДокумент2 страницыAztecas y NayasYelinna Ayling Napuri EchevarriaОценок пока нет
- 7 Paso para Relanzar Tu Marca de ModaДокумент18 страниц7 Paso para Relanzar Tu Marca de ModaShalom GonzalezОценок пока нет
- Dientes Neonatales y NatalesДокумент35 страницDientes Neonatales y Natalescarlos fuentes100% (1)
- Forma Objetiva y Forma Subjetiva - POLIFONÍAДокумент1 страницаForma Objetiva y Forma Subjetiva - POLIFONÍAPablo IrarrázavalОценок пока нет
- 3° Clase Virtual 5°Документ9 страниц3° Clase Virtual 5°silvia salonioОценок пока нет
- Ecologia AplicadaДокумент1 страницаEcologia AplicadaJaneth MirianОценок пока нет
- Climograma Ciudad RealДокумент1 страницаClimograma Ciudad RealluciaОценок пока нет
- Configuración de Sinópticos para La Monitorización y Control de Nivel - Intouch ScadaДокумент16 страницConfiguración de Sinópticos para La Monitorización y Control de Nivel - Intouch ScadaDavid cuasiОценок пока нет
- Manual Poste de RecargaДокумент48 страницManual Poste de RecargaFrancisco Simón FuertesОценок пока нет
- Guía ConductismoДокумент11 страницGuía ConductismoMarta nubia Rivadeneira cortesОценок пока нет
- Monologo de Napoleon de Andres NeumanДокумент4 страницыMonologo de Napoleon de Andres NeumanMoisés GarcíaОценок пока нет
- Conductometria y AperturaДокумент76 страницConductometria y AperturaAlejandra Martinez86% (7)
- Unidad 4. MANTENIMIENTO DEL CULTIVOДокумент7 страницUnidad 4. MANTENIMIENTO DEL CULTIVOAndrés Felipe Giraldo AndradeОценок пока нет
- EVA 2 Corriente ContinúaДокумент9 страницEVA 2 Corriente ContinúaGerman Morera Bedoya100% (1)
- Optimización de Las Necesidades de Agua de La Flor Cortada en Invernadero. Caso Del Cultivo Del ClavelДокумент73 страницыOptimización de Las Necesidades de Agua de La Flor Cortada en Invernadero. Caso Del Cultivo Del ClavelaldairОценок пока нет
- Politica Del SG SST - Compromiso de La Alta DirecciónДокумент3 страницыPolitica Del SG SST - Compromiso de La Alta Direcciónjaime gonzalezОценок пока нет
- Valbol - CatálogoДокумент24 страницыValbol - CatálogoforoОценок пока нет
- Contrato de Arrendamiento de Local ComercialДокумент3 страницыContrato de Arrendamiento de Local ComercialFrancisco Celi CastroОценок пока нет
- Las Nuevas Normas Europeas Armonizadas para Cementos ComunesДокумент23 страницыLas Nuevas Normas Europeas Armonizadas para Cementos ComuneseargmdroОценок пока нет
- Planilla de Turno: Fecha Entrega: 06/11/2021Документ16 страницPlanilla de Turno: Fecha Entrega: 06/11/2021BarbyModestiОценок пока нет
- Producto 3 Npba 280415 Final Inghiver CorregidoДокумент62 страницыProducto 3 Npba 280415 Final Inghiver CorregidoJuan Carlos Grande DescalzoОценок пока нет
- Guia de Logica para Control 1Документ10 страницGuia de Logica para Control 1William L. MontserratОценок пока нет
- Grupo 6-Queso Caquetá PicadoДокумент5 страницGrupo 6-Queso Caquetá PicadoMarlon Stiven Motta ManriqueОценок пока нет
- Rubrica de ExposicionДокумент3 страницыRubrica de ExposicionNorma Cecilia Moreira HerreraОценок пока нет
- Qué Es Chavín de HuántarДокумент10 страницQué Es Chavín de HuántarMj RodrigoОценок пока нет
- Fracciones EquivalentesДокумент2 страницыFracciones EquivalentesBarbara Retamal100% (1)
- Geotecnia Ensayos in SituДокумент35 страницGeotecnia Ensayos in SituSebastian Jimenez RocaОценок пока нет
- La Comprension de LecturaДокумент36 страницLa Comprension de LecturaLorena Mardones RiquelmeОценок пока нет
- Informe Rafael Diaz EsrfДокумент18 страницInforme Rafael Diaz EsrfRAFAEL DIAZ ROJASОценок пока нет