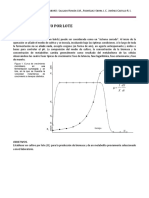
Вам также может понравиться
- Ablandamiento Por Enzimas ExógenasДокумент2 страницыAblandamiento Por Enzimas ExógenasJuan Luis Soto GarciaОценок пока нет
- Examen Final Balance de Materia y EnergiaДокумент2 страницыExamen Final Balance de Materia y EnergiaBRIYID JEHOVANNA TAMI SOLERОценок пока нет
- Manos Seguras Peter SnyderДокумент2 страницыManos Seguras Peter SnyderDeiby RamirezОценок пока нет
- Proceso de Desnatado y Homogenizacion de La LecheДокумент21 страницаProceso de Desnatado y Homogenizacion de La LecheKary Karyta Sanchez0% (1)
- 3 BTL Screening de CepasДокумент10 страниц3 BTL Screening de CepasAlejandro CuetoОценок пока нет
- Caso Práctico Unidad III 2021Документ5 страницCaso Práctico Unidad III 2021paolaОценок пока нет
- Seminario VДокумент10 страницSeminario VTito HuillcaОценок пока нет
- LácteosДокумент9 страницLácteosTephy KuchikiОценок пока нет
- Leche en PolvoДокумент20 страницLeche en PolvoCarla CalvaОценок пока нет
- Practica de Tamizado y MoliendaДокумент14 страницPractica de Tamizado y MoliendaRajid SinningОценок пока нет
- Ejercicios de Alimento CPДокумент37 страницEjercicios de Alimento CPDavid Cabrero GarcíaОценок пока нет
- Determinacion de Solidos Solubles en Jarabes y SalmuerasДокумент6 страницDeterminacion de Solidos Solubles en Jarabes y SalmueraslidiaОценок пока нет
- Transferencia de Masa en Estado EstacionarioДокумент12 страницTransferencia de Masa en Estado EstacionarioWillianPuchoAparicio0% (3)
- Control de Calidad de Conservas VegetalesДокумент10 страницControl de Calidad de Conservas VegetalesGabriela De La Cruz PayanoОценок пока нет
- Elaboración de Nuggets de PolloДокумент11 страницElaboración de Nuggets de PolloIván RichОценок пока нет
- Queso Petit Suisse FINALДокумент35 страницQueso Petit Suisse FINALFernando AlvarezОценок пока нет
- Membranas en Alimentos PDFДокумент7 страницMembranas en Alimentos PDFDavid Reyes LópezОценок пока нет
- Descremado de Leche FrescaДокумент16 страницDescremado de Leche FrescaSony Luis Ramirez HerreraОценок пока нет
- Practica 9 Cultivo Por LoteДокумент4 страницыPractica 9 Cultivo Por LoteAle BernalОценок пока нет
- COMPOSICIÓN DE ALIMENTOS BALANCEADOS - InformeДокумент4 страницыCOMPOSICIÓN DE ALIMENTOS BALANCEADOS - InformeFercho_Pe_a_4388Оценок пока нет
- Tarea 3Документ2 страницыTarea 3Kagney UralesОценок пока нет
- HeladoДокумент8 страницHeladoAnthony CamposОценок пока нет
- Análisis Químico de Los Lípidos PDFДокумент3 страницыAnálisis Químico de Los Lípidos PDFPatricia Belen CarrilloОценок пока нет
- Indice de Solidos en MantequillaДокумент2 страницыIndice de Solidos en MantequillaJudith VegaОценок пока нет
- Ejercicios-Trabajo-Final JVДокумент4 страницыEjercicios-Trabajo-Final JVIsabel OlivaОценок пока нет
- Práctica 1 Difusividad de Un Líquido VolátilДокумент10 страницPráctica 1 Difusividad de Un Líquido VolátilJenny AlvaradoОценок пока нет
- Biorreacor de Columna de BurbujeoДокумент29 страницBiorreacor de Columna de BurbujeoHiara Arenas Lopez0% (1)
- Práctica Propiedades de Las Proteinas de Las Harinas PDFДокумент3 страницыPráctica Propiedades de Las Proteinas de Las Harinas PDFLorena SNОценок пока нет
- Estandarizar LecheДокумент1 страницаEstandarizar LecheOzz Simon100% (1)
- 07 - Extracto de LevaduraДокумент2 страницы07 - Extracto de LevaduraJosé Jesús Gil Ramírez100% (1)
- Punto CrioscopicoДокумент9 страницPunto CrioscopicolidiaОценок пока нет
- Produccion de Amilasas en Cultivo Por LoteДокумент17 страницProduccion de Amilasas en Cultivo Por LotequimikОценок пока нет
- Practica de Dulce de LecheДокумент4 страницыPractica de Dulce de LecheJoel Vlelez JuarezОценок пока нет
- Puntos Críticos de Control Del QuesoДокумент3 страницыPuntos Críticos de Control Del QuesoErika CaceresОценок пока нет
- Quesos Con Propiedades BioactivasДокумент1 страницаQuesos Con Propiedades BioactivasAnarely0% (2)
- BIOSEPARACIONES PRACTICA 7-Saldaña Alejos IbrahimДокумент11 страницBIOSEPARACIONES PRACTICA 7-Saldaña Alejos Ibrahimibrahim saldañaОценок пока нет
- Trabajo Práctico de Balance de Materia y Energía 2017-1Документ4 страницыTrabajo Práctico de Balance de Materia y Energía 2017-1Luis Gutierrez Sanchez0% (1)
- HARINA ModificadoДокумент43 страницыHARINA ModificadoBárbara MoreiraОценок пока нет
- Tercer Parcial BMYEДокумент3 страницыTercer Parcial BMYEAlma GarciaОценок пока нет
- Ejercicios Cultivo Por Lote AlimentadoДокумент3 страницыEjercicios Cultivo Por Lote AlimentadoIvan ParedesОценок пока нет
- Practica 2 FermentaciónДокумент14 страницPractica 2 FermentaciónIris MendezОценок пока нет
- Separacion Por Membranas SubДокумент5 страницSeparacion Por Membranas SubKelly MaritaОценок пока нет
- Reporte de EmulsionesДокумент15 страницReporte de EmulsionesManuel Antonio100% (1)
- Jarabes y SalmuerasДокумент8 страницJarabes y Salmuerasveronica ruiz perezОценок пока нет
- Cuadernos de Cátedra Transf de O2Документ42 страницыCuadernos de Cátedra Transf de O2Rocio CamposОценок пока нет
- Balance de Materia: CajetaДокумент1 страницаBalance de Materia: CajetaIsaac Cruz100% (1)
- Linea de ProduccionДокумент26 страницLinea de ProduccionFabio Leonardo Orozco SernaОценок пока нет
- Guía de Problemas - Balance MateriaДокумент2 страницыGuía de Problemas - Balance Materiamadilo01Оценок пока нет
- SulcaДокумент5 страницSulcaKeny Hans Ramos100% (1)
- Produccion de FrioДокумент12 страницProduccion de FriofrpatriciaОценок пока нет
- Pruebas para La MermeladaДокумент7 страницPruebas para La MermeladaKarlitha DominguezОценок пока нет
- Problemas para Extraccion en ContracorrientefaceДокумент1 страницаProblemas para Extraccion en ContracorrientefaceAlbertoCortésОценок пока нет
- Cuadro Comparativo - Conteo de MicroorganismosДокумент2 страницыCuadro Comparativo - Conteo de MicroorganismosAndrea Yoan Angulo MoralesОценок пока нет
- Practica 6. Elaboracion de QuesoДокумент7 страницPractica 6. Elaboracion de Quesodayana mejiaОценок пока нет
- Informe PastasДокумент4 страницыInforme PastasDidier MelendezОценок пока нет
- Plan HaccpДокумент8 страницPlan HaccpJhon Jairo Pedraza HernandezОценок пока нет
- Balance de EnergiaДокумент13 страницBalance de EnergiaRomario Beltran CoronelОценок пока нет
- Evaporador Triple EfectoДокумент3 страницыEvaporador Triple EfectoHuguiito RcОценок пока нет
- Crecimiento MicrobianoДокумент115 страницCrecimiento MicrobianoLucyRosseCaceresQuispeОценок пока нет
- Modos de Operación de Los FermentadoresДокумент3 страницыModos de Operación de Los FermentadoresRivaJonathanОценок пока нет
- Reco No CimientoДокумент12 страницReco No CimientoYudith Rosaluz Imata CondoriОценок пока нет
- Cierre Perfecto (I2222)Документ4 страницыCierre Perfecto (I2222)Yudith Rosaluz Imata CondoriОценок пока нет
- Remediación Electrocinética JudithДокумент7 страницRemediación Electrocinética JudithYudith Rosaluz Imata CondoriОценок пока нет
- EVAPORADORESДокумент5 страницEVAPORADORESYudith Rosaluz Imata CondoriОценок пока нет
- Mineria Peru y Arequipa-1Документ13 страницMineria Peru y Arequipa-1Yudith Rosaluz Imata CondoriОценок пока нет
- Tesis: Maestro en Ciencia y Tecnología en La Especialidad de Ingeniería AmbientalДокумент99 страницTesis: Maestro en Ciencia y Tecnología en La Especialidad de Ingeniería AmbientalYudith Rosaluz Imata CondoriОценок пока нет
- Filtración 1Документ22 страницыFiltración 1Yudith Rosaluz Imata CondoriОценок пока нет
- Practica 2Документ17 страницPractica 2Yudith Rosaluz Imata CondoriОценок пока нет
- Método de Ranking de FactoresДокумент1 страницаMétodo de Ranking de FactoresYudith Rosaluz Imata CondoriОценок пока нет
- Practicasmicro Indust. 5Документ50 страницPracticasmicro Indust. 5Yudith Rosaluz Imata CondoriОценок пока нет
- Practica 3Документ28 страницPractica 3Yudith Rosaluz Imata CondoriОценок пока нет
- Lixiviacion AvanceДокумент6 страницLixiviacion AvanceYudith Rosaluz Imata CondoriОценок пока нет
- Lab Expo MicroДокумент5 страницLab Expo MicroYudith Rosaluz Imata CondoriОценок пока нет
- Metodologia de CianobacteriasДокумент9 страницMetodologia de CianobacteriasYudith Rosaluz Imata CondoriОценок пока нет
- Elaboración de CremaДокумент6 страницElaboración de CremaYudith Rosaluz Imata Condori100% (2)
- Lab Expo MicroДокумент5 страницLab Expo MicroYudith Rosaluz Imata CondoriОценок пока нет
- Control Del CrecimientoДокумент100 страницControl Del CrecimientoYudith Rosaluz Imata CondoriОценок пока нет
- Nomenclatura IДокумент36 страницNomenclatura ISebastian JaksjakjsОценок пока нет
- Mecanica de Fluidos - Ing ElectromecanicaДокумент9 страницMecanica de Fluidos - Ing ElectromecanicaricardoarianОценок пока нет
- Actividad 4.3. Alteraciones de Las ProteínasДокумент8 страницActividad 4.3. Alteraciones de Las ProteínasBRYAN EDUARDO BRETON DE LA OОценок пока нет
- Reconocimiento de Carbono e Hidrogeno-Nº Lab 1Документ11 страницReconocimiento de Carbono e Hidrogeno-Nº Lab 1YaninaReyes75% (4)
- Unidades de ConcentracionДокумент6 страницUnidades de ConcentracionYulisa Reyes CastilloОценок пока нет
- Fundamento Conceptual6Документ2 страницыFundamento Conceptual6Luigi GuevaraОценок пока нет
- Quimica - Semana 4 - Nomenclatura - Antes y Despues de ClaseДокумент11 страницQuimica - Semana 4 - Nomenclatura - Antes y Despues de Claselusho barrazaОценок пока нет
- Síntesis de Condensación Benzoinica, Bencil y Ácido BencilicoДокумент14 страницSíntesis de Condensación Benzoinica, Bencil y Ácido BencilicoJaime VelásquezОценок пока нет
- Glosario de Ciencias SocialesДокумент5 страницGlosario de Ciencias SocialesGrissel belloОценок пока нет
- LaMedida 2º ESO (Adaptado)Документ3 страницыLaMedida 2º ESO (Adaptado)Julian PopovОценок пока нет
- Tarea 3 - 201424 - 29Документ34 страницыTarea 3 - 201424 - 29Leonardo CastellanosОценок пока нет
- Elipsoide Global Wgs84Документ3 страницыElipsoide Global Wgs84Annie Betzabeth0% (1)
- Tabla Separación de MezclasДокумент2 страницыTabla Separación de MezclasHelena López Alcaraz100% (2)
- Leyes Aire ComprimidoДокумент4 страницыLeyes Aire ComprimidoItalo Hernán Ihpc0% (1)
- Presentación Trabajo de Química y Ciencia Ilustraciones Moléculas Verde TurquesaДокумент12 страницPresentación Trabajo de Química y Ciencia Ilustraciones Moléculas Verde TurquesaNuñez Damian SaraОценок пока нет
- Lab-1 QMC 1206Документ14 страницLab-1 QMC 1206Angie Aliaga Lero100% (1)
- Combustion BiomasaДокумент10 страницCombustion BiomasaAnuar RamosОценок пока нет
- 1.cap V - Diseno de Mezclas Asfalticas en FrioДокумент14 страниц1.cap V - Diseno de Mezclas Asfalticas en FrioJ̶o̶r̶g̶e̶ A̶t̶a̶u̶80% (5)
- Cinemática de MecanismosДокумент7 страницCinemática de MecanismosAlejandro SánchezОценок пока нет
- Aceite de Semilla de Guanabana PDFДокумент9 страницAceite de Semilla de Guanabana PDFRosario PerezОценок пока нет
- Elaboración Del Balance Hídrico y Evaluación de Las Condiciones Sanitarias Del Predio "Marsella" Localizado en El Municipio "El Playón" Departamento de "Santander".Документ14 страницElaboración Del Balance Hídrico y Evaluación de Las Condiciones Sanitarias Del Predio "Marsella" Localizado en El Municipio "El Playón" Departamento de "Santander".Hernan Quispe MendozaОценок пока нет
- Proyecto de Grado UNIPAZДокумент125 страницProyecto de Grado UNIPAZVásquezJuanОценок пока нет
- Rosa de VientoДокумент13 страницRosa de VientoSantos Llicahua HuachacaОценок пока нет
- Tarea 2 - Taller de Conceptos BásicosДокумент15 страницTarea 2 - Taller de Conceptos BásicosAilatan SelivaОценок пока нет
- TTR y Conservación de La MasaДокумент12 страницTTR y Conservación de La MasaLinda RuizОценок пока нет
- Fundamentos Espectroscopia Uv-VisibleДокумент4 страницыFundamentos Espectroscopia Uv-VisibleAvril Correa AparicioОценок пока нет
- Cuestionario Gas 2 by Julio - )Документ2 страницыCuestionario Gas 2 by Julio - )Lionel Villarroel SalazarОценок пока нет
- Laboratorio 5 Versión AlumnoДокумент13 страницLaboratorio 5 Versión AlumnoWilОценок пока нет
- Motor GEET PantoneДокумент42 страницыMotor GEET PantoneLino Lodeiro100% (3)
- Estudio de Pollerias JuliacaДокумент10 страницEstudio de Pollerias JuliacaJheison Favio Quispe BenaventeОценок пока нет