Вам также может понравиться
- El Cerebro de BudaДокумент140 страницEl Cerebro de BudaAndrea100% (12)
- Anatomia Serie RT 8ed PDFДокумент514 страницAnatomia Serie RT 8ed PDFAldo Castillo67% (6)
- Medio Liquido Tema 3Документ5 страницMedio Liquido Tema 3Eliana Churata Rojas100% (1)
- Curso Personal Trainer IFBB Internacional: Fisiología del ejercicioДокумент51 страницаCurso Personal Trainer IFBB Internacional: Fisiología del ejercicioDiego PillacelaОценок пока нет
- Neurofisiología BásicaДокумент258 страницNeurofisiología BásicaWilder Diaz CastilloОценок пока нет
- Uroanalisis Zaragoza PDFДокумент183 страницыUroanalisis Zaragoza PDFRosario LagunaОценок пока нет
- Musa ParadisicaДокумент5 страницMusa ParadisicaVanessaОценок пока нет
- Aislamiento Linfocitos InformeДокумент6 страницAislamiento Linfocitos InformeBelénAlvarezVargasОценок пока нет
- Aprendizaje AceleradoДокумент23 страницыAprendizaje AceleradoEdgar Santiesteban ColladoОценок пока нет
- UntitledДокумент85 страницUntitledMiiguell ReysaОценок пока нет
- Fisiología del sistema nervioso - Organización y funcionesДокумент36 страницFisiología del sistema nervioso - Organización y funcionesmaria puigОценок пока нет
- Bases orgánicas del fenómeno psíquicoДокумент99 страницBases orgánicas del fenómeno psíquicoVazquez Cpsi Silvia80% (5)
- Prunus CerasusДокумент6 страницPrunus CerasusVanessaОценок пока нет
- Prunus CerasusДокумент6 страницPrunus CerasusVanessaОценок пока нет
- Prunus CerasusДокумент6 страницPrunus CerasusVanessaОценок пока нет
- Sustancias Que Producen Los Abrazos en El Cuerpo HumanoДокумент63 страницыSustancias Que Producen Los Abrazos en El Cuerpo Humanoaletintin100% (2)
- Técnicas Histologicas y MicroscopicasДокумент25 страницTécnicas Histologicas y MicroscopicasDoña Tanechka JCОценок пока нет
- Artículo. Maltrato Infantil y Exposición SensibleДокумент10 страницArtículo. Maltrato Infantil y Exposición SensibleBoris Díaz VillegasОценок пока нет
- Hayem A y Hayem BДокумент30 страницHayem A y Hayem BJuan Domingo Davila GilОценок пока нет
- LIQUIDO SEMINAL-1 (1) .PPSXДокумент21 страницаLIQUIDO SEMINAL-1 (1) .PPSXCariito MarquezОценок пока нет
- Informe Laboratorio 2Документ12 страницInforme Laboratorio 2api-25822171450% (2)
- Tema 1 - El Laboratorio de Anatomia PatológicaДокумент11 страницTema 1 - El Laboratorio de Anatomia PatológicachemallinkyОценок пока нет
- CiclosporosisДокумент14 страницCiclosporosisDalia CLОценок пока нет
- El Hombre Como Ser IndescifrableДокумент9 страницEl Hombre Como Ser IndescifrableItzel Herrera100% (1)
- Laboratorio de HematologiaДокумент68 страницLaboratorio de HematologiaJesus Bautisto Huichi JaraОценок пока нет
- Atlas de ParasitologiaДокумент118 страницAtlas de ParasitologiaKaren Alavez0% (1)
- La tinción de Papanicolaou: bases y prácticaДокумент16 страницLa tinción de Papanicolaou: bases y prácticaJeany RoviОценок пока нет
- Mutaciones NADPH oxidasa causan inmunodeficiencia grave combinadaДокумент25 страницMutaciones NADPH oxidasa causan inmunodeficiencia grave combinadaAndrea Escobar100% (1)
- Recuento células hematocitómetroДокумент11 страницRecuento células hematocitómetroMartin CentenoОценок пока нет
- EspermatobiosДокумент2 страницыEspermatobiosAngel PerezОценок пока нет
- PRACTICA No4 LeucocitosДокумент2 страницыPRACTICA No4 LeucocitosGerardo Reyes PulidoОценок пока нет
- S2.Indices Reticulocitos PDFДокумент3 страницыS2.Indices Reticulocitos PDFKenneth Valverde Calderon100% (1)
- PreguntasДокумент7 страницPreguntasDaniela IbarraОценок пока нет
- Informe de MesofilosДокумент9 страницInforme de MesofilosMauro Fidel Mainguez LagosОценок пока нет
- EspermogramaДокумент11 страницEspermogramadragorexОценок пока нет
- Proteinas Doc GДокумент3 страницыProteinas Doc GGuillermo MartinezОценок пока нет
- Causas y tipos de anemiaДокумент36 страницCausas y tipos de anemiaAshura Albaran Evans100% (1)
- Práctica 6 EgoДокумент7 страницPráctica 6 EgoSandy MoralesОценок пока нет
- Presentacion de Bioquimica (Perfiles) JДокумент13 страницPresentacion de Bioquimica (Perfiles) Jjesus lopez100% (1)
- Informe Final Hospitalario CeliacosДокумент14 страницInforme Final Hospitalario CeliacosLineth Salvatierra100% (1)
- Componentes Sanguíneos y CoagulaciónДокумент3 страницыComponentes Sanguíneos y CoagulaciónCitlali Sarai Soto ReyesОценок пока нет
- Trasplantes en Uruguay: Marco legal, ética y estadísticasДокумент16 страницTrasplantes en Uruguay: Marco legal, ética y estadísticasLucia AristimuñoОценок пока нет
- Caso ClinicoДокумент2 страницыCaso ClinicoGeorge ArnaldoОценок пока нет
- Caso ClinicoДокумент9 страницCaso ClinicomilagrosdereОценок пока нет
- Práctica 1Документ2 страницыPráctica 1Alheli BarrientosОценок пока нет
- UtilclinicaДокумент5 страницUtilclinicaLizeth Gonzalez DeviaОценок пока нет
- Antigenos Febriles SPДокумент3 страницыAntigenos Febriles SPRuben Katsura CosmesОценок пока нет
- Caso Clinico HipertiroidismoДокумент12 страницCaso Clinico HipertiroidismoSILVIA KARIN HEREDIA DELGADOОценок пока нет
- Determinación de Bilirrubina y Fosfatasa Alcalina en SueroДокумент21 страницаDeterminación de Bilirrubina y Fosfatasa Alcalina en SueroIsela Castro0% (1)
- 16 Nocardia PDFДокумент37 страниц16 Nocardia PDFHelen FPОценок пока нет
- Examen General de OrinaДокумент9 страницExamen General de OrinaValentina MedellinОценок пока нет
- Liquido Cefalo RaquideoДокумент23 страницыLiquido Cefalo RaquideoAnghelo Bustinza ApazaОценок пока нет
- Tecnicas FijacionДокумент16 страницTecnicas FijacionEiner Jolberth100% (1)
- Evaluacion Analitica Del Metabolismo de Carbohidratos: Tecnólogo Médico Esp. Laboratorio Clínico y Anatomía PatológicaДокумент64 страницыEvaluacion Analitica Del Metabolismo de Carbohidratos: Tecnólogo Médico Esp. Laboratorio Clínico y Anatomía Patológicavalentinatay100% (1)
- BACILLUSДокумент21 страницаBACILLUSLynn FrancisОценок пока нет
- Manual de Prácticas. Realiza Análisis Inmunológicos 2018Документ58 страницManual de Prácticas. Realiza Análisis Inmunológicos 2018Carlos MurilloОценок пока нет
- 02 Guia Separacion de LinfocitosДокумент3 страницы02 Guia Separacion de LinfocitosSophyAraujoОценок пока нет
- UNIDAD VI Anticuerpos Policlonales y MonoclonalesДокумент21 страницаUNIDAD VI Anticuerpos Policlonales y MonoclonalesEmpresa SeguridadОценок пока нет
- Subcultivo de células en monocapa y citometría de flujoДокумент6 страницSubcultivo de células en monocapa y citometría de flujoEduardo LobitoОценок пока нет
- TSH Elisa MonobindДокумент2 страницыTSH Elisa MonobindVini ContrerasОценок пока нет
- COPROCULTIVOДокумент18 страницCOPROCULTIVORychard TextexОценок пока нет
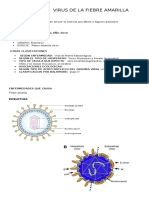
- Fiebre Amarilla Virus: Clasificación y EnfermedadesДокумент3 страницыFiebre Amarilla Virus: Clasificación y EnfermedadesFabian Calderon100% (1)
- APARATO de GOLGIДокумент5 страницAPARATO de GOLGIjose luis sotoОценок пока нет
- Aparato de Golgi, Endocitosis y ExocitosisДокумент20 страницAparato de Golgi, Endocitosis y ExocitosisLuis David Bello RodríguezОценок пока нет
- Aparato de Golgi: estructura y funciones del orgánulo celularДокумент10 страницAparato de Golgi: estructura y funciones del orgánulo celularRodrigo EspinosaОценок пока нет
- Guia Paiep GolgiДокумент11 страницGuia Paiep GolgiSusy FeliОценок пока нет
- Aparato de GolgiДокумент40 страницAparato de GolgiAnette OrtegaОценок пока нет
- Aparato de GolgiДокумент2 страницыAparato de GolgiJhonn Brandon Ventura QuisbertОценок пока нет
- Aparato GolgiДокумент12 страницAparato GolgiSoraida Sanchez Parra0% (1)
- Aparato de GolgiДокумент9 страницAparato de GolgiSebastián TamayoОценок пока нет
- Ensayo Aparato de GolgiДокумент8 страницEnsayo Aparato de GolgiAlina SánchezОценок пока нет
- 7.2 Aparato de GolgiДокумент13 страниц7.2 Aparato de GolgiThelma Zuleiny RamirezОценок пока нет
- Micropropagacion de Solanum TuberosumДокумент1 страницаMicropropagacion de Solanum TuberosumVanessaОценок пока нет
- Efecto de 2,4-DДокумент4 страницыEfecto de 2,4-DVanessaОценок пока нет
- Hormonas VegetalesДокумент1 страницаHormonas VegetalesVanessaОценок пока нет
- TaraxacumДокумент1 страницаTaraxacumVanessaОценок пока нет
- Actividad AntimicrobianaДокумент1 страницаActividad AntimicrobianaVanessaОценок пока нет
- Dieta para UlcerasДокумент1 страницаDieta para UlcerasVanessaОценок пока нет
- Efectos Del Arsénico Sobre La SaludДокумент1 страницаEfectos Del Arsénico Sobre La SaludVanessaОценок пока нет
- HigooДокумент6 страницHigooVanessaОценок пока нет
- Citrus AurantifoliaДокумент7 страницCitrus AurantifoliaVanessaОценок пока нет
- Partenogénesis en Áfidos1Документ1 страницаPartenogénesis en Áfidos1VanessaОценок пока нет
- ArsénicoДокумент2 страницыArsénicoVanessaОценок пока нет
- Partenogénesis 1Документ1 страницаPartenogénesis 1VanessaОценок пока нет
- Efectos Del Arsénico Sobre La SaludДокумент1 страницаEfectos Del Arsénico Sobre La SaludVanessaОценок пока нет
- Ciudadania e IdentidadДокумент2 страницыCiudadania e IdentidadVanessaОценок пока нет
- TERMORREGULACIÓNДокумент2 страницыTERMORREGULACIÓNVanessaОценок пока нет
- Iseminario MolecularДокумент8 страницIseminario MolecularVanessaОценок пока нет
- Expo Molecular Polimorfismo UrosДокумент15 страницExpo Molecular Polimorfismo UrosVanessaОценок пока нет
- Análisis MultivariadoДокумент2 страницыAnálisis MultivariadoVanessaОценок пока нет
- Ciudadania e IdentidadДокумент2 страницыCiudadania e IdentidadVanessaОценок пока нет
- ORQUIDEASДокумент6 страницORQUIDEASVanessaОценок пока нет
- Objetiv OsДокумент2 страницыObjetiv OsVanessaОценок пока нет
- Cuestionario 8Документ5 страницCuestionario 8VanessaОценок пока нет
- PROTEINAS SNARE Y GTPasas DE TRANSPORTE 1Документ4 страницыPROTEINAS SNARE Y GTPasas DE TRANSPORTE 1VanessaОценок пока нет
- La Principal Función Del Cerebro Es Mantener Vivo Al Organismo para Que Interactúe Con El EntornoДокумент6 страницLa Principal Función Del Cerebro Es Mantener Vivo Al Organismo para Que Interactúe Con El EntornoVanessaОценок пока нет
- Demostracion de Algunas Acciones Del Sistema Nervioso Simpatico y Parasimpatico en ConejosДокумент6 страницDemostracion de Algunas Acciones Del Sistema Nervioso Simpatico y Parasimpatico en ConejosVanessaОценок пока нет
- Glosario Funcional del Sistema NerviosoДокумент65 страницGlosario Funcional del Sistema NerviosoAna ToraОценок пока нет
- Resumen de Fisiología y Física Biológica 2019Документ180 страницResumen de Fisiología y Física Biológica 2019Candela SarmientoОценок пока нет
- Sistema nervioso fisiologíaДокумент15 страницSistema nervioso fisiologíaNeibeer Diaz100% (2)
- LABORATORIO No 4 MicroanatomiaДокумент4 страницыLABORATORIO No 4 MicroanatomiaMichiru LainzОценок пока нет
- 5-Sistema SomatosensorialДокумент13 страниц5-Sistema SomatosensorialJavier FernandezОценок пока нет
- Sistema nervioso y sus funcionesДокумент4 страницыSistema nervioso y sus funcionesGerard Sorto100% (1)
- Neurona y SinapsisДокумент16 страницNeurona y SinapsisYuliana Valverde BarreiroОценок пока нет
- Tipos de Neuronas - Características y Funciones PDFДокумент16 страницTipos de Neuronas - Características y Funciones PDFmuchachaОценок пока нет
- Taller de Biologia 2 Sistema NerviosoДокумент13 страницTaller de Biologia 2 Sistema NerviosoMaria Paula Baron100% (5)
- Diferencias Y Similitudes Entre Potencial Graduado Y Potencias de AcciónДокумент15 страницDiferencias Y Similitudes Entre Potencial Graduado Y Potencias de Acciónflopii melloОценок пока нет
- Neurona-Georgina MéndezДокумент5 страницNeurona-Georgina MéndezgmendezzОценок пока нет
- Bioelectricidad de tejidos excitablesДокумент5 страницBioelectricidad de tejidos excitablesAdrian ParedesОценок пока нет
- Neurodesarrollo InfantilДокумент19 страницNeurodesarrollo InfantilElizabeth RandoОценок пока нет
- II Unofidad Morfisiologia III Completa Samuel Reyes-2Документ252 страницыII Unofidad Morfisiologia III Completa Samuel Reyes-2Gerald EduardooОценок пока нет
- AtlasДокумент8 страницAtlasdessterОценок пока нет
- Guia Examen de HistologiaДокумент25 страницGuia Examen de HistologiaYorgelis Paola Querales MendezОценок пока нет
- Apunte C - Intro SN IIДокумент22 страницыApunte C - Intro SN IIcarolina cerveraОценок пока нет
- Clase 4. Tejido NerviosoДокумент37 страницClase 4. Tejido NerviosoRuben Galvarino Tapia RamosОценок пока нет
- Psicobiologia Unidad 1 - Ciclo de La Tarea 1-Estructura Del Trabajo A Entregar (1) 2Документ9 страницPsicobiologia Unidad 1 - Ciclo de La Tarea 1-Estructura Del Trabajo A Entregar (1) 2Elizabeth Mosquera Martinez100% (1)
- Sesión de Aprendizaje 18Документ40 страницSesión de Aprendizaje 18JHONATAN ACOSTA SEOPAОценок пока нет
- ANATOMÍAДокумент10 страницANATOMÍAMonica NancyОценок пока нет