Вам также может понравиться
- ArqueobacteriasДокумент2 страницыArqueobacteriasEnrique Luna Guerrero100% (4)
- Extremofilos PDFДокумент46 страницExtremofilos PDFJosue VillotaОценок пока нет
- TEMA N°1 y 2 ARQUEAS Y VIRUS PDFДокумент7 страницTEMA N°1 y 2 ARQUEAS Y VIRUS PDFLuis Valencia HerenciaОценок пока нет
- TEMA N°5 y 6 ARQUEAS Y VIRUSДокумент9 страницTEMA N°5 y 6 ARQUEAS Y VIRUSJoal2000Оценок пока нет
- TEMA N°5 y 6 ARQUEAS Y VIRUSДокумент9 страницTEMA N°5 y 6 ARQUEAS Y VIRUSMelany Ramirez JavierОценок пока нет
- Album de DominiosДокумент13 страницAlbum de DominiosJefferson De LeónОценок пока нет
- Dominio Archaea 201888Документ8 страницDominio Archaea 201888Corales JoseОценок пока нет
- Reino ArchaeaДокумент2 страницыReino ArchaeaGianella L-MoralesОценок пока нет
- EuryaarchaeotaДокумент3 страницыEuryaarchaeotaDidier ChambillaОценок пока нет
- ARCHAEДокумент4 страницыARCHAEazmobileguiniОценок пока нет
- Eur y Archaeo TaДокумент4 страницыEur y Archaeo Ta6622010007 CAMILA MARIA RODRIGUEZ VARGAS ESTUDIANTE ACTIVOОценок пока нет
- Arqueo BacteriasДокумент3 страницыArqueo BacteriasJosé Carlos CcqОценок пока нет
- Dominio ArcheaДокумент11 страницDominio ArcheaROSA PORTERO RODRIGUEZОценок пока нет
- Arbol FilogeneticoДокумент40 страницArbol FilogeneticoFabián MoralesОценок пока нет
- Bbic U2 A2 JobgДокумент6 страницBbic U2 A2 JobgYhoss Zahyd Bdillo RomeroОценок пока нет
- Ar Que o BacteriasДокумент3 страницыAr Que o BacteriasAnonymous KFBlpx6SrfОценок пока нет
- Reino ArqueobacteriaДокумент4 страницыReino Arqueobacteriajohann espinolaОценок пока нет
- Lectura de BacteriasДокумент2 страницыLectura de BacteriasMargarita CayoОценок пока нет
- Actividad de Investigación - MicrobiologíaДокумент10 страницActividad de Investigación - Microbiologíaserna monseОценок пока нет
- Bacterias y ArqueobacteriasДокумент3 страницыBacterias y ArqueobacteriasDaniela Rodriguez Davila100% (1)
- Ar Que Ob Act TeriasДокумент8 страницAr Que Ob Act TeriasAlejandro OlmosОценок пока нет
- HalofilosДокумент15 страницHalofilosCristian Javier Bustamante De La HozОценок пока нет
- ARQUEOBACTERIASДокумент5 страницARQUEOBACTERIASDavid Huayana HurtadoОценок пока нет
- Archa BacteriasДокумент4 страницыArcha BacteriasElizabeth Bonilla ReaОценок пока нет
- Reino ArchaeaДокумент2 страницыReino ArchaeaLuz Alejandra VasquezОценок пока нет
- ArqueasДокумент39 страницArqueasJosé Larragaña Osuna100% (3)
- Dominio ArchaeaДокумент8 страницDominio ArchaeaJoseph PaucarОценок пока нет
- Relacion de Microorg. Con El Medio Ambiente-1 - 254Документ43 страницыRelacion de Microorg. Con El Medio Ambiente-1 - 254Karol Laura LucasОценок пока нет
- FeriaДокумент13 страницFeriaMiguel CoronaОценок пока нет
- EXTREMOFILOSДокумент14 страницEXTREMOFILOSYinny OrtizОценок пока нет
- Son Las Bacterias Primitivas Que Habitan Lugares de Alta Concentración SalinaДокумент10 страницSon Las Bacterias Primitivas Que Habitan Lugares de Alta Concentración SalinaKarinitah Levano CruzОценок пока нет
- ArqueobacteriasДокумент2 страницыArqueobacteriaspedro meyer alvaron cristobalОценок пока нет
- Las ArqueobacteriasДокумент2 страницыLas ArqueobacteriasBLACRUZH14Оценок пока нет
- ArcheaДокумент10 страницArcheaJose David Morales DОценок пока нет
- ArqueobacteriaДокумент31 страницаArqueobacteriaGoncast GhiobertiОценок пока нет
- Extremofilos 1Документ15 страницExtremofilos 1Yinny OrtizОценок пока нет
- U3g Extremofilos 20267 PDFДокумент36 страницU3g Extremofilos 20267 PDFHector Sierra100% (1)
- 10 MicrofósilesДокумент17 страниц10 MicrofósilesAnonymous dtTVDeОценок пока нет
- Presentación1CYT ARQUEAДокумент6 страницPresentación1CYT ARQUEANoelia AnicetoОценок пока нет
- Ultimo Informe TeoricoДокумент7 страницUltimo Informe TeoricoLis VeraОценок пока нет
- ARQUEOBACTERIASДокумент9 страницARQUEOBACTERIASFernando Pinzón BurgaОценок пока нет
- Los Tres DominiosДокумент36 страницLos Tres DominiosValeria Calderón100% (1)
- Ar Que o BacteriaДокумент10 страницAr Que o BacteriaYael Jimenez LopezОценок пока нет
- ArqueobacteriasДокумент7 страницArqueobacteriasLis VeraОценок пока нет
- ARQUEOBACTERIASДокумент18 страницARQUEOBACTERIASJesus Alain Perez RuizОценок пока нет
- Los Tres Dominios de La VidaДокумент6 страницLos Tres Dominios de La Vidalizbeth pichardoОценок пока нет
- Microrganismo en Ambiente Extremos 2017Документ19 страницMicrorganismo en Ambiente Extremos 2017RichardSarmientoPalominoОценок пока нет
- Reino MoneraДокумент16 страницReino MoneraJhamylet OrdoñezОценок пока нет
- Niveles de Organización y Clasificación de Los Seres Vivos Equipo 2Документ19 страницNiveles de Organización y Clasificación de Los Seres Vivos Equipo 2Josué Osvaldo Enciso GarcíaОценок пока нет
- Reino BacteriaДокумент3 страницыReino Bacteriakatherine Jasmin melgar CastilloОценок пока нет
- Árbol Filogenetico UniversalДокумент12 страницÁrbol Filogenetico UniversalMariana Vilta100% (1)
- ARQUEOBACTERIASДокумент9 страницARQUEOBACTERIASElizabeth PahuachoОценок пока нет
- ArqueasДокумент3 страницыArqueascarlos cuervoОценок пока нет
- Ejer Cici OsДокумент22 страницыEjer Cici Oscatherine garcia mirandaОценок пока нет
- Invertebrados de Brusca Capitulo 1Документ21 страницаInvertebrados de Brusca Capitulo 1jenieduОценок пока нет
- Micro InformeДокумент13 страницMicro InformeNancy ObrienОценок пока нет
- Reino ArchaebacteriaДокумент2 страницыReino ArchaebacteriaSilvia ParroneОценок пока нет
- GRUPO 6-Procesos Metabolicos de Los Hiperthermophile (MICRO AMBIENTAL)Документ11 страницGRUPO 6-Procesos Metabolicos de Los Hiperthermophile (MICRO AMBIENTAL)Jorge Yaya JuarezОценок пока нет
- Las pirañas: Anatomía, alimentación, reproducción, enfermedades, cría en acuarioОт EverandLas pirañas: Anatomía, alimentación, reproducción, enfermedades, cría en acuarioОценок пока нет
- Iii Bimestre - V Unidad - BiologiaДокумент14 страницIii Bimestre - V Unidad - BiologiaFelipe RecobaОценок пока нет
- Clase 4 - 5to Primaria - QuimicaДокумент4 страницыClase 4 - 5to Primaria - QuimicaFelipe RecobaОценок пока нет
- Clase 3 - 5to Primaria - QuímicaДокумент26 страницClase 3 - 5to Primaria - QuímicaFelipe RecobaОценок пока нет
- Clase 4 - 5to Primaria - BiologíaДокумент6 страницClase 4 - 5to Primaria - BiologíaFelipe RecobaОценок пока нет
- Clase 1 - 5to Primaria - BiologíaДокумент2 страницыClase 1 - 5to Primaria - BiologíaFelipe RecobaОценок пока нет
- 4 El-Continente-AsiaticoДокумент4 страницы4 El-Continente-AsiaticoFelipe RecobaОценок пока нет
- Clase 1 - 5to Primaria - QuimicaДокумент2 страницыClase 1 - 5to Primaria - QuimicaFelipe RecobaОценок пока нет
- 3 Historia Civilismo PDFДокумент2 страницы3 Historia Civilismo PDFFelipe RecobaОценок пока нет
- 3 El-Continente-EuropeoДокумент4 страницы3 El-Continente-EuropeoFelipe RecobaОценок пока нет
- Nomenclatura Inorganica TeoriaДокумент7 страницNomenclatura Inorganica TeoriaFelipe RecobaОценок пока нет
- Pretextos Del ConflictoДокумент1 страницаPretextos Del ConflictoFelipe RecobaОценок пока нет
- 2 Historia La Guerra Con España PDFДокумент3 страницы2 Historia La Guerra Con España PDFFelipe Recoba100% (1)
- Exposicion de La Guerra de España - Historia.Документ12 страницExposicion de La Guerra de España - Historia.Felipe RecobaОценок пока нет
- CotizacionДокумент2 страницыCotizacionFelipe RecobaОценок пока нет
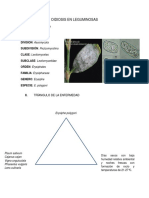
- Oidiosis en LeguminosasДокумент2 страницыOidiosis en LeguminosasFelipe RecobaОценок пока нет
- Unidad 1 de Biologia BasicaДокумент42 страницыUnidad 1 de Biologia BasicaMaria Prieto DiazОценок пока нет
- Glosario BacteriologíaДокумент16 страницGlosario BacteriologíaViridiana SahianОценок пока нет
- Clasificacion de Los Seres Vivos TallerДокумент7 страницClasificacion de Los Seres Vivos TallerFelipe AyaОценок пока нет
- Solucionario 2 Jeg LenguajeДокумент11 страницSolucionario 2 Jeg LenguajeSalvador ToledoОценок пока нет
- Lectura Celula Teoria PDFДокумент8 страницLectura Celula Teoria PDFAleja MancholaОценок пока нет
- Secundaria 1 Trim 2Документ286 страницSecundaria 1 Trim 2MARIMONTEBEL100% (3)
- Sistemas PrecelularesДокумент11 страницSistemas PrecelularesKarlicka Chavez0% (1)
- Célula ProcariotaДокумент33 страницыCélula ProcariotaIsa GonzalezОценок пока нет
- Definición de Reino MoneraДокумент3 страницыDefinición de Reino MoneraJimena MartinОценок пока нет
- ArcheobacteriasДокумент9 страницArcheobacteriasMarioVelka0% (1)
- Células Eucariotas y ProcariotasДокумент5 страницCélulas Eucariotas y ProcariotasINTERNET R y GОценок пока нет
- Conceptos Sobre MicrobiologíaДокумент125 страницConceptos Sobre MicrobiologíaLuis Andres Chavez Lopez100% (1)
- Evolución Del CitoesqueletoДокумент11 страницEvolución Del CitoesqueletoDiego ArriagadaОценок пока нет
- Biologia - Tema 1Документ53 страницыBiologia - Tema 1Lianna TrujillanoОценок пока нет
- 20201-Celulas Procariotas y EucariotasДокумент38 страниц20201-Celulas Procariotas y EucariotasRossy EstradaОценок пока нет
- Trabajo Encargado CyaДокумент23 страницыTrabajo Encargado CyaFrank Luis Jimenez MamaniОценок пока нет
- ArqueasДокумент17 страницArqueasGIOVANNI DAVID BRIGANTE LUNAОценок пока нет
- Expo de Biologia 1Документ10 страницExpo de Biologia 1Karla AparicioОценок пока нет
- Las Categorías TaxonómicasДокумент7 страницLas Categorías TaxonómicasRoberto TorresОценок пока нет
- Arqueas y La Ultra EstructuraДокумент5 страницArqueas y La Ultra EstructuraLiz VillarОценок пока нет
- Preguntas Guía Sobre El Texto de MadiganДокумент7 страницPreguntas Guía Sobre El Texto de MadiganValenthinaCamargoОценок пока нет
- Microbiologia, Parasitologia e InmunologiaДокумент77 страницMicrobiologia, Parasitologia e Inmunologiacarolina cuevas rosОценок пока нет
- Bergeys Manual of Determinative BacterioДокумент11 страницBergeys Manual of Determinative Bacteriomayra yainОценок пока нет
- Reino ArchaeaДокумент9 страницReino ArchaeaCeleste ValladaresОценок пока нет
- Examen de Microbiologia ResueltoДокумент5 страницExamen de Microbiologia ResueltoGiovanna GarcíaОценок пока нет
- Exposicion de Crecimiento 6.3 y 6.4Документ11 страницExposicion de Crecimiento 6.3 y 6.4Lizbeth AlcivarОценок пока нет
- Guerrero y Berlanga, 2005Документ9 страницGuerrero y Berlanga, 2005Noe Hernandez TorrejonОценок пока нет
- Tema 6Документ36 страницTema 6Santiago QuezadaОценок пока нет
- Factores Intrinsecos y Extrinsecos Que Afectan Al Crecimiento MicrobianoДокумент27 страницFactores Intrinsecos y Extrinsecos Que Afectan Al Crecimiento MicrobianoBartz KrauserОценок пока нет
- Reino MoneraДокумент11 страницReino MoneraVanessa Almeida FontalvoОценок пока нет