Вам также может понравиться
- Level of Consciousness Refers To A Dimension That Varies From Coma at One Extreme, ThroughДокумент3 страницыLevel of Consciousness Refers To A Dimension That Varies From Coma at One Extreme, Throughkristinush1991100% (1)
- @MBS MedicalBooksStore 2018 The PDFДокумент1 061 страница@MBS MedicalBooksStore 2018 The PDFDhiya Muthiah Gaffari100% (6)
- Medical Mnemonics-Internal MedicineДокумент38 страницMedical Mnemonics-Internal MedicineRaman Goyal100% (1)
- Brain Chip ReportДокумент30 страницBrain Chip Reportsrikanthkalemla100% (3)
- Miracles in The BrainДокумент10 страницMiracles in The BrainIVAN ALVAREZОценок пока нет
- CENTRAL ANALGETICS Analgesics Chemistry Pharmacology of Drugs Daniel Lednicer 1982Документ236 страницCENTRAL ANALGETICS Analgesics Chemistry Pharmacology of Drugs Daniel Lednicer 1982BASKAR.LОценок пока нет
- 421 - NeurobiologyДокумент38 страниц421 - NeurobiologymelodyfathiОценок пока нет
- 0262011816.MIT Press - Sunny Y. Auyang - Life, Mind in Everyday & Science, Cognitive - May.2009Документ526 страниц0262011816.MIT Press - Sunny Y. Auyang - Life, Mind in Everyday & Science, Cognitive - May.2009Branko NikolicОценок пока нет
- Emotion and The Human Brain: Antonio R. DamasioДокумент6 страницEmotion and The Human Brain: Antonio R. DamasioBruno XavierОценок пока нет
- Immortality of ConsciousnessДокумент8 страницImmortality of Consciousnessmd91101Оценок пока нет
- Brain Facts Book 2018 High Res PDFДокумент140 страницBrain Facts Book 2018 High Res PDFMadeley Yáñez NavarroОценок пока нет
- Neurophilosophy and The Healthy MindДокумент6 страницNeurophilosophy and The Healthy MindNortonMentalHealthОценок пока нет
- Baron Psychology Notes PDFДокумент78 страницBaron Psychology Notes PDFRamya MeesalaОценок пока нет
- Sleep Physiology: DR Herman Mulijadi MS, SPKPДокумент41 страницаSleep Physiology: DR Herman Mulijadi MS, SPKPCandice LavigneОценок пока нет
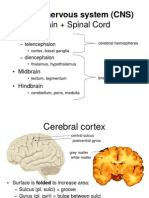
- Brain + Spinal Cord: Central Nervous System (CNS)Документ18 страницBrain + Spinal Cord: Central Nervous System (CNS)Anna Marthea V.100% (4)
- Sensory System-2Документ35 страницSensory System-2samar yousif mohamedОценок пока нет
- Quantum Mechanics of 'Conscious Energy'Документ29 страницQuantum Mechanics of 'Conscious Energy'SYED ISMYL MAHMOOD RIZVIОценок пока нет
- What Are The Parts of The Brain and Their FunctionsДокумент2 страницыWhat Are The Parts of The Brain and Their FunctionskimuyyyyОценок пока нет
- TheДокумент680 страницTheMónica GarzaОценок пока нет
- How Our Brain Is Thinking? AbstractДокумент7 страницHow Our Brain Is Thinking? AbstractsyedmoulaliОценок пока нет
- 2020 - 10 - 29 - Εγκεφαλικά Ημισφαίρια και Αγγειακά Σύνδρομα - ΜήτσιαςДокумент67 страниц2020 - 10 - 29 - Εγκεφαλικά Ημισφαίρια και Αγγειακά Σύνδρομα - ΜήτσιαςΖέτα ΤσίρκαОценок пока нет
- Clinical Aspects of Sensory Motor IntegrationДокумент350 страницClinical Aspects of Sensory Motor IntegrationAlexandra AiftimieОценок пока нет
- Top 10 Unsolved Mysteries of The BrainДокумент8 страницTop 10 Unsolved Mysteries of The BrainLokesh KhuranaОценок пока нет
- Erro Ahmad Surya 1611900027 Academic Writing AДокумент5 страницErro Ahmad Surya 1611900027 Academic Writing AErro SuryaОценок пока нет
- Consciousness Origins of PDFДокумент6 страницConsciousness Origins of PDF11111Оценок пока нет
- Nerves of PerceptionДокумент18 страницNerves of PerceptionJOHNATIELОценок пока нет
- The Brain - An Without A Conductor: OrchestraДокумент3 страницыThe Brain - An Without A Conductor: OrchestralauranadarОценок пока нет
- Chapter 6 - States of ConsciousnessДокумент56 страницChapter 6 - States of ConsciousnessakshitaОценок пока нет
- Cerebro y MenteДокумент5 страницCerebro y MenteCarlos DaboinОценок пока нет
- El Surgimiento de La ConcienciaДокумент6 страницEl Surgimiento de La ConcienciaRafael AitaОценок пока нет
- Consciousness As Integrated InformationДокумент27 страницConsciousness As Integrated InformationjackОценок пока нет
- Exploring Neuroscience 14-20Документ7 страницExploring Neuroscience 14-20Patricia Susckind100% (1)
- Consciousness As Integrated Information: A Provisional ManifestoДокумент27 страницConsciousness As Integrated Information: A Provisional ManifestothyagosmesmeОценок пока нет
- Mashour 2013Документ8 страницMashour 2013Carolina LuditzaОценок пока нет
- Philosophy and The Sciences: What Is Consciousness? (Part II)Документ6 страницPhilosophy and The Sciences: What Is Consciousness? (Part II)jcvoscribОценок пока нет
- The Sleeping BrainДокумент11 страницThe Sleeping BrainGabriela Cristina García FacioОценок пока нет
- Massimini PBR Coma Science 2009Документ14 страницMassimini PBR Coma Science 2009hamletquironОценок пока нет
- Exam 4 Study Guide Fall 2017: I. Cognition & Consciousness: The Nature of ConsciousnessДокумент7 страницExam 4 Study Guide Fall 2017: I. Cognition & Consciousness: The Nature of ConsciousnessAnonymous C6dXawSОценок пока нет
- Consciência - o Problema Real - Anil K SethДокумент8 страницConsciência - o Problema Real - Anil K SethMarcos Paulo RamosОценок пока нет
- Cog ReviewerДокумент4 страницыCog ReviewerShanna Delos SantosОценок пока нет
- Thought Paper 2 - Nancy DongДокумент6 страницThought Paper 2 - Nancy DongNancy D.Оценок пока нет
- Onscious Awareness and Behaviour What Distinguishes Conscious Experience From Unconscious ProcessesДокумент20 страницOnscious Awareness and Behaviour What Distinguishes Conscious Experience From Unconscious Processesj.h. FERNANDEZОценок пока нет
- Neuroplasticity - or Brain PlasticityДокумент2 страницыNeuroplasticity - or Brain Plasticityjack stewartОценок пока нет
- Barry. Perception and Visual Communication TheoryДокумент16 страницBarry. Perception and Visual Communication TheoryKosta Hoekstra50% (2)
- 2005 - Baars - Global Workspace Theory of Consciousness - Toward A Cognitive Neuroscience of Human ExperienceДокумент9 страниц2005 - Baars - Global Workspace Theory of Consciousness - Toward A Cognitive Neuroscience of Human ExperiencefachavalОценок пока нет
- Sleep, Emotions, and Visceral Control: I. N. Pigarev and M. L. PigarevaДокумент12 страницSleep, Emotions, and Visceral Control: I. N. Pigarev and M. L. PigarevaMariaОценок пока нет
- Edelman, Gerald - Neural DarwinismДокумент8 страницEdelman, Gerald - Neural DarwinismcdmemfgambОценок пока нет
- Chapter 1 ReadingДокумент4 страницыChapter 1 ReadingYousef Ahmad2Оценок пока нет
- Editorial: J.Neurol Sci (Turk)Документ4 страницыEditorial: J.Neurol Sci (Turk)Nezih OktarОценок пока нет
- The Brain: Learning ObjectivesДокумент16 страницThe Brain: Learning Objectivessaswati biswalОценок пока нет
- Neuroanatomyof AttentionДокумент21 страницаNeuroanatomyof AttentionPs. Jean CastroОценок пока нет
- Module 10.11Документ7 страницModule 10.11Michaella PagaduraОценок пока нет
- Neural Plasticity and Consciousness (Figures Are Missing in This Version) Susan HurleyДокумент35 страницNeural Plasticity and Consciousness (Figures Are Missing in This Version) Susan HurleyA_WretchОценок пока нет
- Chapter 3 NeuroscienceДокумент6 страницChapter 3 NeuroscienceskateboardinglivОценок пока нет
- Lit ReviewДокумент12 страницLit Reviewapi-318508032Оценок пока нет
- Brain and MindДокумент26 страницBrain and MindPeter M.K. ChanОценок пока нет
- F. Crick C. KochДокумент13 страницF. Crick C. KochDacio NascimentoОценок пока нет
- The Brain's Cerebral Cortex. The Lobes of The Cerebral Cortex and New TechnologiesДокумент12 страницThe Brain's Cerebral Cortex. The Lobes of The Cerebral Cortex and New TechnologiesAgustin PortilloОценок пока нет
- Ijas 2011Документ24 страницыIjas 2011neurodeidaОценок пока нет
- ITP-States of ConsciousnessДокумент17 страницITP-States of ConsciousnesspopiscanzОценок пока нет
- Connecting Consciousness: The Role of Large Scale Brain Networks in ConsciousnessДокумент33 страницыConnecting Consciousness: The Role of Large Scale Brain Networks in ConsciousnessHariОценок пока нет
- 54 The Brain From Inside Out 15: Chapter 1. e ProblemДокумент20 страниц54 The Brain From Inside Out 15: Chapter 1. e ProblemjuannnnОценок пока нет
- Cognitive P. Chapter 2Документ4 страницыCognitive P. Chapter 2May AloОценок пока нет
- The Brain - NobaДокумент21 страницаThe Brain - NobaMarОценок пока нет
- Brainy Mind GREGORYДокумент3 страницыBrainy Mind GREGORYtrasibulo1Оценок пока нет
- ConsciousnessДокумент4 страницыConsciousnessEzekiel ScherzenbachОценок пока нет
- Chapter 3 Biological Foundations of BehaviourДокумент20 страницChapter 3 Biological Foundations of BehaviourAlex LiОценок пока нет
- Kuliah Medula SpinalisДокумент85 страницKuliah Medula SpinalisMuhamad Thohar ArifinОценок пока нет
- Peter Wolf - History of Epilepsy - Nosological Concepts and ClassificationДокумент9 страницPeter Wolf - History of Epilepsy - Nosological Concepts and Classificationgoo99Оценок пока нет
- Anatomical Studies of Brainstem-Evoked Antinociceptive ControlsДокумент194 страницыAnatomical Studies of Brainstem-Evoked Antinociceptive ControlsnervefiberОценок пока нет
- Diencephalon PhysiologyДокумент36 страницDiencephalon PhysiologyNatnael0% (1)
- First Few Seconds For FlowДокумент18 страницFirst Few Seconds For FlowMateiDCОценок пока нет
- 1 - Somatosensory System IДокумент5 страниц1 - Somatosensory System ITony DawaОценок пока нет
- Bases Neurales de La Violencia HumanaДокумент9 страницBases Neurales de La Violencia HumanaJoji von SchwedlerОценок пока нет
- Dementia: Role of MRI: Updated VersionДокумент30 страницDementia: Role of MRI: Updated VersionAnonymous 61SwB8efОценок пока нет
- Stroke Syndromes: Hemiparesis (Faciobrachiocrural Weakness)Документ7 страницStroke Syndromes: Hemiparesis (Faciobrachiocrural Weakness)eko aОценок пока нет
- Insular CortexДокумент7 страницInsular Cortexelbueno21Оценок пока нет
- Pusher Behaviour A Critical Review of Controversial IssuesДокумент11 страницPusher Behaviour A Critical Review of Controversial Issues林芝生Оценок пока нет
- CH 7 The Nervous System NotesДокумент21 страницаCH 7 The Nervous System Notesbiswa217Оценок пока нет
- NREM and REM SleepДокумент17 страницNREM and REM SleepJulius Czarces AgustinОценок пока нет
- The Sleep-Deprived Human BrainДокумент15 страницThe Sleep-Deprived Human BrainHector AngelОценок пока нет
- Introduction A Guide To NeurolocalizationДокумент7 страницIntroduction A Guide To NeurolocalizationTizazu BayihОценок пока нет
- Neuroanatomy Text and Atlas 4 e Inkling Chapter Enhanced Ebook Neuroanatomy Text Atlas Martin 4th Edition Ebook PDFДокумент62 страницыNeuroanatomy Text and Atlas 4 e Inkling Chapter Enhanced Ebook Neuroanatomy Text Atlas Martin 4th Edition Ebook PDFgerald.mims287100% (39)
- Journal of Child Neurology 4Документ7 страницJournal of Child Neurology 4NEUROLAB ESCALÓNОценок пока нет
- Limbic System NeuroscienceДокумент82 страницыLimbic System Neuroscience250187155Оценок пока нет