Вам также может понравиться
- Stem Cell Anthology: From Stem Cell Biology, Tissue Engineering, Cloning, Regenerative Medicine and BiologyОт EverandStem Cell Anthology: From Stem Cell Biology, Tissue Engineering, Cloning, Regenerative Medicine and BiologyОценок пока нет
- Embryonic Stem Cells in Development and Regenerative MedicineДокумент15 страницEmbryonic Stem Cells in Development and Regenerative Medicineallyssagardoce08Оценок пока нет
- OLLI Stem CellДокумент46 страницOLLI Stem CellthevapesandtechltdОценок пока нет
- Stem CellsДокумент3 страницыStem CellsLhoNa ﭢ ArpilledaОценок пока нет
- Embryonic Stem Cell Properties and UseДокумент6 страницEmbryonic Stem Cell Properties and UseRaj Kumar SoniОценок пока нет
- 2012 PLoS One - Selective Development of Myogenic Mesenchymal Cells From Human Embryonic and Induced Pluripotent Stem CellsДокумент9 страниц2012 PLoS One - Selective Development of Myogenic Mesenchymal Cells From Human Embryonic and Induced Pluripotent Stem Cellstomo-awayaОценок пока нет
- Stem Cells - A Recent Approach: Dr. D. Sudarsanam ProfessorДокумент55 страницStem Cells - A Recent Approach: Dr. D. Sudarsanam ProfessorPoonkodi TpОценок пока нет
- Stem CellДокумент4 страницыStem CellZeth VargasОценок пока нет
- Regenerative Medicine 2006Документ106 страницRegenerative Medicine 2006Mdo3Оценок пока нет
- Pluripotent Nontumorigenic Multilineage Differentiating Stress Enduring Cells (Muse Cells) : A Seven-Year RetrospectiveДокумент9 страницPluripotent Nontumorigenic Multilineage Differentiating Stress Enduring Cells (Muse Cells) : A Seven-Year RetrospectiveMaximiliano MichellОценок пока нет
- Embryonic Stem Cells: Review ArticleДокумент6 страницEmbryonic Stem Cells: Review ArticleFabLab EIUОценок пока нет
- Scaffolds For Stem CellsДокумент8 страницScaffolds For Stem Cellscubed3Оценок пока нет
- Stem CellsДокумент29 страницStem CellsZahraaОценок пока нет
- Stem Cell Transplantation As A Mode Of...Документ6 страницStem Cell Transplantation As A Mode Of...Becky GingerОценок пока нет
- Cardiac Stem Cell Therapy: An Overview: AKM M Islam, AAS Majumder, F Doza, MM Rahman, H JesminДокумент15 страницCardiac Stem Cell Therapy: An Overview: AKM M Islam, AAS Majumder, F Doza, MM Rahman, H JesminNavojit ChowdhuryОценок пока нет
- Stem CellДокумент9 страницStem CellDulce UribeОценок пока нет
- Stem Cells in The ClinicДокумент8 страницStem Cells in The ClinicKamal AdeyemiОценок пока нет
- (Review) (Biomolecular Concepts) Kazuyuki Ohbo Et Shin-Ichi Tomizawa-2015Документ9 страниц(Review) (Biomolecular Concepts) Kazuyuki Ohbo Et Shin-Ichi Tomizawa-2015Priscilla FreschuОценок пока нет
- Stem Cell Therapy: A Novel & Futuristic Treatment Modality For Disaster InjuriesДокумент17 страницStem Cell Therapy: A Novel & Futuristic Treatment Modality For Disaster InjuriesChiboyKrenzОценок пока нет
- Adipose Tissue Derived Stem Cells For Regeneration and Differentiation Into InsulinProducing CellsДокумент5 страницAdipose Tissue Derived Stem Cells For Regeneration and Differentiation Into InsulinProducing CellsRodolflopezОценок пока нет
- Summer 2007 Workshop in Biology and Multimedia For High School TeachersДокумент45 страницSummer 2007 Workshop in Biology and Multimedia For High School TeachersmarysssssssssОценок пока нет
- The Difference of Stem Cell ProductДокумент12 страницThe Difference of Stem Cell ProductIsmail Tidak Ada KepanjanganОценок пока нет
- Stem Cell: Cellula PrecursoriaДокумент26 страницStem Cell: Cellula PrecursoriaYhan Brotamonte BoneoОценок пока нет
- StemcellsДокумент41 страницаStemcellsN CH MadhusudhanОценок пока нет
- A Chemical Approach To Stem-Cell Biology and Regenerative MedicineДокумент7 страницA Chemical Approach To Stem-Cell Biology and Regenerative MedicinenembutalОценок пока нет
- Farin 2009Документ25 страницFarin 2009Laura Duarte RojasОценок пока нет
- Stem CellДокумент8 страницStem CellHanumat SinghОценок пока нет
- 1 B. Teknologi Sel Punca Terapi HatiДокумент12 страниц1 B. Teknologi Sel Punca Terapi HatiarisОценок пока нет
- Stem Cell: Navigation SearchДокумент14 страницStem Cell: Navigation Searchanon-589691Оценок пока нет
- Stem Cell: Mihai - Cristian BAGHINA Seria A Grupa 2Документ9 страницStem Cell: Mihai - Cristian BAGHINA Seria A Grupa 2Mihai NycromОценок пока нет
- The Concept of Radiation-Enhanced Stem Cell Differentiation: Research ArticleДокумент8 страницThe Concept of Radiation-Enhanced Stem Cell Differentiation: Research ArticledwinugrohojuandaОценок пока нет
- What Are Stem Cells and Where Do They Come FromДокумент4 страницыWhat Are Stem Cells and Where Do They Come FromjaniellegbОценок пока нет
- Stem Cell TherapyДокумент26 страницStem Cell TherapymfspongebobОценок пока нет
- Potency of Various Types of Stem Cells and Their Transplantation 2157 7633.1000115Документ6 страницPotency of Various Types of Stem Cells and Their Transplantation 2157 7633.1000115VILEOLAGOLDОценок пока нет
- VIT University, Vellore: Animal Physiology and Biotechnology Assignment OnДокумент10 страницVIT University, Vellore: Animal Physiology and Biotechnology Assignment OnSwapnil TiwariОценок пока нет
- Introduction To Stem CellsДокумент32 страницыIntroduction To Stem CellsAtif Amin Baig100% (2)
- Stem Cell Proposal Final 2003.Документ13 страницStem Cell Proposal Final 2003.Harshal GohilОценок пока нет
- Potential of Induced Pluripotent Stem Cells For Use in Gene TherapyДокумент16 страницPotential of Induced Pluripotent Stem Cells For Use in Gene TherapyAnnaaaaaОценок пока нет
- Tissue Renewal, Regeneration and RepairДокумент50 страницTissue Renewal, Regeneration and RepairNurul AfiahОценок пока нет
- Therapeutic Potential of Embryonic Stem CellsДокумент11 страницTherapeutic Potential of Embryonic Stem CellsMike LopezОценок пока нет
- Stem Cells - A Revolution in Medicine - PDДокумент20 страницStem Cells - A Revolution in Medicine - PDTatiane Cruz da SilvaОценок пока нет
- Stem Cells: Vitro and in VivoДокумент14 страницStem Cells: Vitro and in VivoSamsonОценок пока нет
- The Use of Embryonic Stem CellsFNДокумент5 страницThe Use of Embryonic Stem CellsFNPeter KimathiОценок пока нет
- Adult Stem CellsДокумент9 страницAdult Stem CellsPedro Henrique100% (1)
- Stem Cell ResearchДокумент4 страницыStem Cell Researchpianomaster82Оценок пока нет
- Stem Cells in Gynecology 2012 American Journal of Obstetrics and GynecologyДокумент8 страницStem Cells in Gynecology 2012 American Journal of Obstetrics and GynecologyfujimeisterОценок пока нет
- Stem-Cell CultureДокумент14 страницStem-Cell CultureArnob DriptoОценок пока нет
- Stem Cells in Regenerative Medicine and Therapeutic PotentialДокумент12 страницStem Cells in Regenerative Medicine and Therapeutic PotentialIJAR JOURNALОценок пока нет
- Stem Cells Thesis PDFДокумент4 страницыStem Cells Thesis PDFHelpWithWritingPapersPittsburgh100% (2)
- Collas 03 Teaching Cells New TricksДокумент8 страницCollas 03 Teaching Cells New TricksFrancesco ValsegaОценок пока нет
- ISSCR 11 StemCellFactBrch FNLДокумент8 страницISSCR 11 StemCellFactBrch FNLНваневу СильванусОценок пока нет
- Applications in Molecular BiotechnologyДокумент8 страницApplications in Molecular BiotechnologyRaj BanalaОценок пока нет
- Stem CellsДокумент58 страницStem CellsRakesh Kumar100% (2)
- InTech-Stem Cells Ethical and Religious Issues PDFДокумент17 страницInTech-Stem Cells Ethical and Religious Issues PDFFionna PohanОценок пока нет
- Regenerative Medicine: Presented By: Darshita Sharma Roll No: 17/IBT/046Документ22 страницыRegenerative Medicine: Presented By: Darshita Sharma Roll No: 17/IBT/046Darshita VashishthaОценок пока нет
- Generation of Patient Specific Stem Cells: A Human Model SystemДокумент23 страницыGeneration of Patient Specific Stem Cells: A Human Model Systemkj185Оценок пока нет
- Stem Cell Application and UsesДокумент12 страницStem Cell Application and UsesSwarnapaliliyanageОценок пока нет
- The Anatomical Record Part A Discoveries in Molecular Cellular and Evolutionary Biology - 2003 - Gerecht NirДокумент8 страницThe Anatomical Record Part A Discoveries in Molecular Cellular and Evolutionary Biology - 2003 - Gerecht Nirstevenburrow06Оценок пока нет
- Advances in Reprogramming Somatic Cells To InducedДокумент21 страницаAdvances in Reprogramming Somatic Cells To InducedNatalia MoysésОценок пока нет
- Reviewer ThermecДокумент1 страницаReviewer ThermecKhristiam AlvarezОценок пока нет
- Nozzle Loads Piping Stresses and The Effect of Piping On EquipmentДокумент40 страницNozzle Loads Piping Stresses and The Effect of Piping On Equipmentcaesar00100% (6)
- WOMBAT InnovationДокумент57 страницWOMBAT InnovationKhristiam AlvarezОценок пока нет
- Multiphase Pumps Leistritz Article Oct2011Документ2 страницыMultiphase Pumps Leistritz Article Oct2011Khristiam AlvarezОценок пока нет
- Equivalent Length of Pipe PDFДокумент3 страницыEquivalent Length of Pipe PDFSSudhakarОценок пока нет
- DavincinotesДокумент1 страницаDavincinotesping_remullaОценок пока нет
- 1992 Field Test Poseidon MPPДокумент10 страниц1992 Field Test Poseidon MPPKhristiam AlvarezОценок пока нет
- Whats Wrong With MurderДокумент21 страницаWhats Wrong With MurderKhristiam AlvarezОценок пока нет
- WT Article Ozone 5 04Документ3 страницыWT Article Ozone 5 04Khristiam AlvarezОценок пока нет
- If Your Clothes Aren't Already Made Out of Plastic, They Will Be - QuartzДокумент6 страницIf Your Clothes Aren't Already Made Out of Plastic, They Will Be - QuartzKhristiam AlvarezОценок пока нет
- Seismic Design of Earth Retaining StructuresДокумент9 страницSeismic Design of Earth Retaining StructuresMehul JethvaОценок пока нет
- Energy in Figures 2012Документ58 страницEnergy in Figures 2012Khristiam AlvarezОценок пока нет
- Como Fazer Um GroutДокумент4 страницыComo Fazer Um Grouthudson.lopes6380Оценок пока нет
- FLIR - Thermal Imaging GuidebookДокумент48 страницFLIR - Thermal Imaging Guidebookdaniel_silabanОценок пока нет
- 6S TrainingДокумент40 страниц6S Trainingayaskant751001100% (2)
- Axial Load Frequencies BeamsДокумент33 страницыAxial Load Frequencies BeamsKhristiam AlvarezОценок пока нет
- Practical Consideration For Pump Baseplates, Grouting and FoundationДокумент6 страницPractical Consideration For Pump Baseplates, Grouting and FoundationВладимир СмирновОценок пока нет
- Yamaha - Sound Reinforcement HandbookДокумент431 страницаYamaha - Sound Reinforcement HandbookEduardo Rios PortuguezОценок пока нет
- Timeline Project Planning: 12 MonthsДокумент12 страницTimeline Project Planning: 12 MonthsKhristiam AlvarezОценок пока нет
- CFD Simulation of Multi Phase Twin Screw PumpДокумент56 страницCFD Simulation of Multi Phase Twin Screw PumpSoumendu Banik100% (1)
- Sleeve Vs Antifriction Bearings Selection of The Optimal BearingДокумент13 страницSleeve Vs Antifriction Bearings Selection of The Optimal BearingKamal Arab0% (1)
- MEK4450 FlowAssurance Pensum-2Документ58 страницMEK4450 FlowAssurance Pensum-2hiyeonОценок пока нет
- 2006 THESIS Wet-Gas Compression in TSMPДокумент115 страниц2006 THESIS Wet-Gas Compression in TSMPKhristiam AlvarezОценок пока нет
- Agni Dagdha. Group-T Patients Were Treated With Indigenous Drugs and Group-C PatientsДокумент14 страницAgni Dagdha. Group-T Patients Were Treated With Indigenous Drugs and Group-C PatientsKrishnaОценок пока нет
- Improving Performance With Feedback, Rewards, and Positive ReinforcementДокумент40 страницImproving Performance With Feedback, Rewards, and Positive ReinforcementCalla YeeОценок пока нет
- Potassium: Why Is This Medication Prescribed?Документ6 страницPotassium: Why Is This Medication Prescribed?Anonymous mFvEL96SОценок пока нет
- The Voice: C T V BДокумент12 страницThe Voice: C T V BJulia GavrashenkoОценок пока нет
- Articles For TOEFLДокумент8 страницArticles For TOEFLEpp Jb BernatОценок пока нет
- Managing Insomnia Lesson 3Документ20 страницManaging Insomnia Lesson 3taurus_europeОценок пока нет
- Anesthesia Coding GuidelinesДокумент71 страницаAnesthesia Coding GuidelinesAjay IyerОценок пока нет
- Stroke Statistics 2018Документ46 страницStroke Statistics 2018Aghnia PutriОценок пока нет
- Video Game AddictionДокумент29 страницVideo Game AddictionNikunj Agrawal0% (1)
- SGD Lupus Nephritis NotesДокумент6 страницSGD Lupus Nephritis NotesfrnksusОценок пока нет
- Primitive ReflexesДокумент6 страницPrimitive ReflexesMarikit2012100% (1)
- Pediatric OncologyДокумент57 страницPediatric Oncologyjoycechicago100% (2)
- Decompensated HFДокумент16 страницDecompensated HFJessica GarletsОценок пока нет
- The Neurobiology of Personality DisordersДокумент10 страницThe Neurobiology of Personality Disordersfan-a-tykОценок пока нет
- EMQs For Medical Students Volume 3 2eДокумент16 страницEMQs For Medical Students Volume 3 2ePasTestBooks50% (2)
- Waller, G. Et Al (2020)Документ10 страницWaller, G. Et Al (2020)Carlos Emilt Zarco Montes de OcaОценок пока нет
- End-Ball Manual - EnGДокумент20 страницEnd-Ball Manual - EnGJunyong LeeОценок пока нет
- ComputationsДокумент10 страницComputationsFaye Nervanna Alecha AlferezОценок пока нет
- Children's Community Nursing Team - SOP 14 - Oxygen Therapy For ChildrenДокумент8 страницChildren's Community Nursing Team - SOP 14 - Oxygen Therapy For ChildrenSajanan TrimedОценок пока нет
- Abg Specimen Collection Test RequisitionДокумент7 страницAbg Specimen Collection Test RequisitionVangie OriginesОценок пока нет
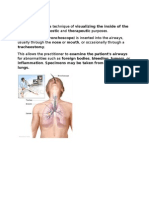
- BronchosДокумент12 страницBronchosAhmad ShyoukhОценок пока нет
- Busen Proposal1Документ47 страницBusen Proposal1Prativa NeupaneОценок пока нет
- Hotspot RecordДокумент3 страницыHotspot RecordIoanna SachaОценок пока нет
- Biblio MethДокумент103 страницыBiblio MethRichard DodsonОценок пока нет
- Guillain Barre SyndromeДокумент8 страницGuillain Barre SyndromeStephanie AureliaОценок пока нет
- Hydrocollator Pack ApplicationДокумент4 страницыHydrocollator Pack ApplicationSreeraj S R100% (1)
- Melodic Intonation TherapyДокумент6 страницMelodic Intonation Therapyrocio100% (1)
- SL Report-278574812Документ2 страницыSL Report-278574812SiddharthОценок пока нет
- Is Your Home Harming You?Документ4 страницыIs Your Home Harming You?Iris Brooks100% (2)