Вам также может понравиться
- Repair Manual 100 - 200 - 300 Series Automatic Pack Land Camera - March 1970Документ81 страницаRepair Manual 100 - 200 - 300 Series Automatic Pack Land Camera - March 1970Elizabeth Perry Muderick100% (4)
- 5 Tips For Better Low Light Images: PhotzyДокумент15 страниц5 Tips For Better Low Light Images: PhotzymrpiracyОценок пока нет
- 09 - Congenital SyndromesДокумент76 страниц09 - Congenital SyndromesROHIT100% (1)
- L-3WarriorSystems 2013 ProductGuide DigitalEditionДокумент1 страницаL-3WarriorSystems 2013 ProductGuide DigitalEditionAlexjohn2009100% (1)
- OTC103002 OptiX BWS 1600G System Description ISSUE1.0Документ37 страницOTC103002 OptiX BWS 1600G System Description ISSUE1.0rohit00722Оценок пока нет
- Infrared IR Sensor Circuit Diagram and Working PrincipleДокумент7 страницInfrared IR Sensor Circuit Diagram and Working PrincipleKirz SerranoОценок пока нет
- Scan Plan Paut-Mc-02-Corregido PDFДокумент5 страницScan Plan Paut-Mc-02-Corregido PDFCarlos TijerinaОценок пока нет
- Optical OptogeneticsДокумент22 страницыOptical OptogeneticshisenknОценок пока нет
- Body MRI Sequences: A Conceptual Framework: Applied Radiology January 2012Документ7 страницBody MRI Sequences: A Conceptual Framework: Applied Radiology January 2012Akanda RabbiОценок пока нет
- Coene N 2011Документ7 страницCoene N 2011Estereotaxia BrasilОценок пока нет
- High-Resolution Sonography of The Peripheral Nervous System - A Review of The LiteratureДокумент10 страницHigh-Resolution Sonography of The Peripheral Nervous System - A Review of The LiteratureJP ChenОценок пока нет
- Mapping Neuronal DiversityДокумент2 страницыMapping Neuronal DiversityJohn MistryОценок пока нет
- Tam Et Al-2015-Journal of NeurochemistryДокумент16 страницTam Et Al-2015-Journal of NeurochemistryAMОценок пока нет
- Imaging Fast Electrical Activity in The Brain With Electrical Impedance TomographyДокумент10 страницImaging Fast Electrical Activity in The Brain With Electrical Impedance TomographyKhởi Trịnh TuấnОценок пока нет
- Coherent OscillationsДокумент10 страницCoherent OscillationsAlejandraОценок пока нет
- Berning 2012Документ1 страницаBerning 2012László SágiОценок пока нет
- A Practical Guide To Photoacoustic Tomography in The Life SciencesДокумент12 страницA Practical Guide To Photoacoustic Tomography in The Life SciencesyuyiipОценок пока нет
- Sequence Optimization For MRI Acoustic NoiseДокумент9 страницSequence Optimization For MRI Acoustic NoiseArsy ZygomaticОценок пока нет
- Imaging Techniques For The Lumbar Spine: Conventional Radiology, Computed Tomography Magnetic Resonance ImagingДокумент23 страницыImaging Techniques For The Lumbar Spine: Conventional Radiology, Computed Tomography Magnetic Resonance ImagingfilibusterosОценок пока нет
- Fnsyn 12 00036Документ26 страницFnsyn 12 00036Epes KadizadeОценок пока нет
- Zha Burst Imaging Review ISUM 1999Документ6 страницZha Burst Imaging Review ISUM 1999LEPING ZHAОценок пока нет
- The Use of Laser Doppler Vibrometry Doppler Principle For Middle Ear Research and DiagnosisДокумент8 страницThe Use of Laser Doppler Vibrometry Doppler Principle For Middle Ear Research and DiagnosisHoria MocanuОценок пока нет
- Nanohole Biosensor-Origin and Application As Multiplex Biosensing PlatformДокумент3 страницыNanohole Biosensor-Origin and Application As Multiplex Biosensing PlatformShibsekhar RoyОценок пока нет
- Mot Artifact Removal fNIRS MeintjesДокумент11 страницMot Artifact Removal fNIRS MeintjesAlex WongОценок пока нет
- Artifacts and Pitfalls US Part 2Документ15 страницArtifacts and Pitfalls US Part 2Abhishek LonikarОценок пока нет
- Basic Principles of QEEGДокумент7 страницBasic Principles of QEEGdietapОценок пока нет
- tmp9198 TMPДокумент13 страницtmp9198 TMPFrontiersОценок пока нет
- tmp719B TMPДокумент13 страницtmp719B TMPFrontiersОценок пока нет
- Fabian I 2007Документ3 страницыFabian I 2007meripe sureshОценок пока нет
- 2003 Changes in Median Nerve Somatosensory Transmission and Motor OutputДокумент12 страниц2003 Changes in Median Nerve Somatosensory Transmission and Motor OutputchooОценок пока нет
- Hearing Research: Hisayuki Ojima, Kunio MurakamiДокумент8 страницHearing Research: Hisayuki Ojima, Kunio MurakamiFrontiersОценок пока нет
- Supre Re Soul Tion ConceptsДокумент5 страницSupre Re Soul Tion Conceptsalka mehraОценок пока нет
- Noise and Vestibular Perception of Passive Self-Motion: Francesco Lacquaniti, Barbara La Scaleia and Myrka ZagoДокумент19 страницNoise and Vestibular Perception of Passive Self-Motion: Francesco Lacquaniti, Barbara La Scaleia and Myrka ZagoRahma watiОценок пока нет
- Chapter28 The Kindling PhenomenonДокумент13 страницChapter28 The Kindling Phenomenonwenshancheng2023Оценок пока нет
- A New Diagnostic Tool For Detecting Dental Caries and Tooth Wear: Ultrasonic SystemДокумент4 страницыA New Diagnostic Tool For Detecting Dental Caries and Tooth Wear: Ultrasonic SystempoojaОценок пока нет
- MRI-Based Attenuation Correction For PET/MRI Using Ultrashort Echo Time SequencesДокумент8 страницMRI-Based Attenuation Correction For PET/MRI Using Ultrashort Echo Time SequencesOmarah AbdalqaderОценок пока нет
- Basic Principles of Diagnostic UltrasonographyДокумент5 страницBasic Principles of Diagnostic UltrasonographyAgung KusastiОценок пока нет
- 1 s2.0 S0167876015001865 MainДокумент11 страниц1 s2.0 S0167876015001865 Mainsantosh kumarОценок пока нет
- Resonancia Magnetica en Animales de CompañíaДокумент11 страницResonancia Magnetica en Animales de CompañíaEnrique A. Vega NuñezОценок пока нет
- Deán-Ben 2011 Phys. Med. Biol. 56 6129Документ21 страницаDeán-Ben 2011 Phys. Med. Biol. 56 6129kostismavrakisОценок пока нет
- Informatics in Medicine Unlocked: SciencedirectДокумент15 страницInformatics in Medicine Unlocked: SciencedirectIon CorbuОценок пока нет
- Le Bihan 98Документ3 страницыLe Bihan 98Sameh RaoufОценок пока нет
- Hearing Research: Lingling Cai, Glenna Stomackin, Nicholas M. Perez, Xiaohui Lin, Timothy T. Jung, Wei DongДокумент10 страницHearing Research: Lingling Cai, Glenna Stomackin, Nicholas M. Perez, Xiaohui Lin, Timothy T. Jung, Wei DongJasmy FfОценок пока нет
- Tugas Keperwatan Dasar Kelompok 3Документ54 страницыTugas Keperwatan Dasar Kelompok 3Tolak HarisОценок пока нет
- 19-Variational Dynamics of Bose-Einstein Condensates in Deep Optical LatticesДокумент11 страниц19-Variational Dynamics of Bose-Einstein Condensates in Deep Optical LatticesIsrael PeñaОценок пока нет
- Probe Oscillation Shear Elastography (PROSE)Документ9 страницProbe Oscillation Shear Elastography (PROSE)oscar.atondoОценок пока нет
- Stimulated Emission Depletion (STED) Nanoscopy of A Fluorescent Protein-Labeled Organelle Inside A Living CellДокумент6 страницStimulated Emission Depletion (STED) Nanoscopy of A Fluorescent Protein-Labeled Organelle Inside A Living CellAMОценок пока нет
- Convention Paper: A Low-End Device To Convert EEG Waves To MusicДокумент7 страницConvention Paper: A Low-End Device To Convert EEG Waves To MusicAdrian Attard TrevisanОценок пока нет
- A New Ultrasound Method For Determining The AcousticДокумент10 страницA New Ultrasound Method For Determining The AcousticMale RamìrezОценок пока нет
- Lecture 1-1Документ19 страницLecture 1-1Masood ChoudharyОценок пока нет
- Inhibición y Curvas de Sintonía en NCV - FullДокумент14 страницInhibición y Curvas de Sintonía en NCV - FullSilvio SánchezОценок пока нет
- Isochronous Space-Time Wave Packets: Corresponding Author: Raddy@creol - Ucf.eduДокумент5 страницIsochronous Space-Time Wave Packets: Corresponding Author: Raddy@creol - Ucf.eduGerové InvestmentsОценок пока нет
- Graded Activation of The Intrinsic Laryngeal Muscles For Vocal Fold PosturingДокумент7 страницGraded Activation of The Intrinsic Laryngeal Muscles For Vocal Fold PosturingDaniel AlonsoОценок пока нет
- A Carbon-Fiber Electrode Array For Long-Term Neural RecordingДокумент22 страницыA Carbon-Fiber Electrode Array For Long-Term Neural RecordingDavide RicciОценок пока нет
- Me471 6Документ3 страницыMe471 6ehteshamalhanifОценок пока нет
- TSLIM Imaging and A Morphometric Analysis of The Mouse SP - 2011 - Hearing ReseaДокумент9 страницTSLIM Imaging and A Morphometric Analysis of The Mouse SP - 2011 - Hearing ReseaRoger Fernando Abril DiazОценок пока нет
- Zundorf 2015 Dual Pathway Auditory SystemДокумент10 страницZundorf 2015 Dual Pathway Auditory SystemRicardo Jose De LeonОценок пока нет
- Foltz 2012Документ19 страницFoltz 2012alejandro.david1642Оценок пока нет
- Influence of Detection Conditions On Near-Field Optical ImagingДокумент11 страницInfluence of Detection Conditions On Near-Field Optical ImagingClaudio BiaginiОценок пока нет
- Neuroimage: Justin Ales, Thom Carney, Stanley A. KleinДокумент9 страницNeuroimage: Justin Ales, Thom Carney, Stanley A. KleinnikadonОценок пока нет
- Radtke-Schuller2016 Article BrainAtlasOfTheMongolianGerbilДокумент272 страницыRadtke-Schuller2016 Article BrainAtlasOfTheMongolianGerbilCharlotte NordenОценок пока нет
- Genomic RemodellingДокумент2 страницыGenomic RemodellingDenys Quispe AlcocerОценок пока нет
- Fernandez-Miranda JC. 2007 - Three-Dimensional Microsurgical and Tractographic Anatomy of The White Matter..Документ40 страницFernandez-Miranda JC. 2007 - Three-Dimensional Microsurgical and Tractographic Anatomy of The White Matter..Beyza ZeyrekОценок пока нет
- Vaina 1990Документ17 страницVaina 1990Monica VazОценок пока нет
- OCT Imaging in Glaucoma: A guide for practitionersОт EverandOCT Imaging in Glaucoma: A guide for practitionersKi Ho ParkОценок пока нет
- G Onshor 1976Документ34 страницыG Onshor 1976ROHITОценок пока нет
- TB TreatДокумент47 страницTB TreatROHITОценок пока нет
- 92 FullДокумент5 страниц92 FullROHITОценок пока нет
- The Visual White Matter: The Application of Diffusion MRI and Fiber Tractography To Vision ScienceДокумент50 страницThe Visual White Matter: The Application of Diffusion MRI and Fiber Tractography To Vision ScienceROHITОценок пока нет
- Neuroimage: Clinical: ArticleinfoДокумент8 страницNeuroimage: Clinical: ArticleinfoROHITОценок пока нет
- 2015 6 jns1546Документ7 страниц2015 6 jns1546ROHITОценок пока нет
- ECG & Heart Sounds: Student HandoutДокумент7 страницECG & Heart Sounds: Student HandoutROHITОценок пока нет
- Atrial Fibrillation and Atrial FlutterДокумент4 страницыAtrial Fibrillation and Atrial FlutterROHITОценок пока нет
- Vision Research: Oliver Braddick, Janette AtkinsonДокумент22 страницыVision Research: Oliver Braddick, Janette AtkinsonROHITОценок пока нет
- AphsДокумент4 страницыAphsROHITОценок пока нет
- Illd Lp78 Pl78 Dill EngДокумент10 страницIlld Lp78 Pl78 Dill EngROHITОценок пока нет
- Reference BooksДокумент5 страницReference BooksROHITОценок пока нет
- III 123.fullДокумент8 страницIII 123.fullROHITОценок пока нет
- MCB 150: Molecular Immunology: Spring 2007 Prof. Ellen Robey Prof. Robert Beatty Prof. Laurent CoscoyДокумент70 страницMCB 150: Molecular Immunology: Spring 2007 Prof. Ellen Robey Prof. Robert Beatty Prof. Laurent CoscoyROHITОценок пока нет
- Introduction Immunology 000Документ123 страницыIntroduction Immunology 000ROHITОценок пока нет
- SN Chez 1984Документ6 страницSN Chez 1984ROHITОценок пока нет
- Sezary SyndromeДокумент13 страницSezary SyndromeROHITОценок пока нет
- AlifДокумент2 страницыAlifROHITОценок пока нет
- I. General Pathology: Athology Exam Questions FOR Students OF DentistryДокумент8 страницI. General Pathology: Athology Exam Questions FOR Students OF DentistryROHITОценок пока нет
- Linear-to-Circular Polarization Conversion Using MetasurfaceДокумент6 страницLinear-to-Circular Polarization Conversion Using MetasurfacenaveenОценок пока нет
- Scientech 2502Документ2 страницыScientech 2502Rishi KeshОценок пока нет
- TelescopeДокумент3 страницыTelescopeTijani Basit AbiodunОценок пока нет
- Microscope and Its ClassificationДокумент26 страницMicroscope and Its Classificationsharmahemlata603Оценок пока нет
- L16.2 OPHTHA Ocular Motility (Nov1622)Документ5 страницL16.2 OPHTHA Ocular Motility (Nov1622)Erald PaderangaОценок пока нет
- PHYSICAL-SCIENCE-week 7 MODULE-11Документ32 страницыPHYSICAL-SCIENCE-week 7 MODULE-11EMОценок пока нет
- Laporan Praktikum IДокумент6 страницLaporan Praktikum IIrfan fajrulОценок пока нет
- 01 5 CF BFДокумент357 страниц01 5 CF BFblaiОценок пока нет
- Darkroom and Film Processing Handout RAD TECH AДокумент2 страницыDarkroom and Film Processing Handout RAD TECH ADomingo McKenzie May P.Оценок пока нет
- THE Microscope: ObjectivesДокумент8 страницTHE Microscope: ObjectiveskimОценок пока нет
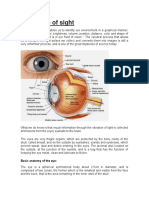
- The Sense of SightДокумент7 страницThe Sense of SightRosy SantanaОценок пока нет
- 3D Display - PROJECTOR - BARCO LED FL40Документ2 страницы3D Display - PROJECTOR - BARCO LED FL40Correo DerespaldoОценок пока нет
- Bragg'S Law, and Diffractometer: by Ayesha SiddiqaДокумент11 страницBragg'S Law, and Diffractometer: by Ayesha SiddiqaAyesha SiddiqaОценок пока нет
- Gallery of Diverse Cells: Microscope Lab By: Emma KimuraДокумент10 страницGallery of Diverse Cells: Microscope Lab By: Emma Kimuraapi-346991843Оценок пока нет
- CCD Calibrations and Noise Considerations - Karen KwitterДокумент19 страницCCD Calibrations and Noise Considerations - Karen KwitterMechtatel HrenovОценок пока нет
- Lecture 04 Chain Surveying-1Документ38 страницLecture 04 Chain Surveying-1Mutahir KhattakОценок пока нет
- Chapter 4 - Optical Fiber ConnectionДокумент39 страницChapter 4 - Optical Fiber ConnectionFaizal EngintechОценок пока нет
- Aperture, Shutter Speed, IsoДокумент2 страницыAperture, Shutter Speed, IsoAndrei RaduОценок пока нет
- Vision & Visual Acuity (Smart Optometry)Документ14 страницVision & Visual Acuity (Smart Optometry)Disha Varun MamniyaОценок пока нет
- Physics Investigatory ProjectДокумент17 страницPhysics Investigatory Projectyatishsingh1975Оценок пока нет
- BS-M101 MergedДокумент11 страницBS-M101 MergedArkaprabha GhoshОценок пока нет
- TE-440 Optical Fiber Communication - Mid-Term - v4Документ3 страницыTE-440 Optical Fiber Communication - Mid-Term - v4Muhammad ImranОценок пока нет
- Dokumen - Tips Nikon d3 User ManualДокумент471 страницаDokumen - Tips Nikon d3 User ManualJohnОценок пока нет
- Chemistry: Unit 2: Assignment 1 Analytical Methods and Separation TechniquesДокумент30 страницChemistry: Unit 2: Assignment 1 Analytical Methods and Separation TechniquesLorri - Ann Lamont100% (1)