Вам также может понравиться
- Study PCOS ObesityДокумент9 страницStudy PCOS ObesitydfdfffffОценок пока нет
- Molecular Mechanism of Obesity-Induced Metabolic' Tissue RemodelingДокумент6 страницMolecular Mechanism of Obesity-Induced Metabolic' Tissue RemodelingKanaОценок пока нет
- Adipose Tissue Expandability, Lipotoxicity and The Metabolic SyndromeДокумент12 страницAdipose Tissue Expandability, Lipotoxicity and The Metabolic SyndromePsicología del SerОценок пока нет
- Ectopic Fat and Insulin Resistance: Pathophysiology and Effect of Diet and Lifestyle InterventionsДокумент19 страницEctopic Fat and Insulin Resistance: Pathophysiology and Effect of Diet and Lifestyle InterventionsIlknur Okyay KellyОценок пока нет
- Ouchi, 2015Документ30 страницOuchi, 2015wanda oktariaОценок пока нет
- Cell Biology of Fat StorageДокумент5 страницCell Biology of Fat StorageMauricio VidalОценок пока нет
- JURNAL - Jaringan AdiposaДокумент10 страницJURNAL - Jaringan AdiposaPramesvara NaoriОценок пока нет
- HHS Public Access: Metabolic Adaptation and Maladaptation in Adipose TissueДокумент27 страницHHS Public Access: Metabolic Adaptation and Maladaptation in Adipose TissueJulia SCОценок пока нет
- ##1. Adipose-Tissue Plasticity in Health and DiseaseДокумент28 страниц##1. Adipose-Tissue Plasticity in Health and Diseasejonsmth552Оценок пока нет
- Importance of The Microenvironment and Mechanosensing in Adipose Tissue BiologyДокумент28 страницImportance of The Microenvironment and Mechanosensing in Adipose Tissue BiologyAnonymous HUY0yRexYfОценок пока нет
- Fat Tissue Aging and Cellular SenescenceДокумент18 страницFat Tissue Aging and Cellular SenescenceFilda de LimaОценок пока нет
- Adipose Tissue Plasticity From WAT To BAT and in BetweenДокумент12 страницAdipose Tissue Plasticity From WAT To BAT and in Betweenjonsmth552Оценок пока нет
- Review Article: Adipocytokines Novel Link Between Inflammation and Vascular Function?Документ24 страницыReview Article: Adipocytokines Novel Link Between Inflammation and Vascular Function?dlupu_30Оценок пока нет
- Artigo 7Документ7 страницArtigo 7daisyscholliersОценок пока нет
- 202307戴宛霖Inflamed Adipose TissueДокумент13 страниц202307戴宛霖Inflamed Adipose Tissuemiguel contrerasОценок пока нет
- Obesida ArticuloДокумент5 страницObesida ArticuloZangre AzulОценок пока нет
- Nutrients 14 01509Документ15 страницNutrients 14 01509Miguel AlejandroОценок пока нет
- Adipocyte Differentiation - From Fibroblast To Endocrine Cell2001Документ8 страницAdipocyte Differentiation - From Fibroblast To Endocrine Cell2001Reza AzghadiОценок пока нет
- Role of The Adipocyte in MetabolismДокумент30 страницRole of The Adipocyte in MetabolismAna Gariza SolanoОценок пока нет
- 2018overnutrition Ectopic Lipid and The Metabolic SyndromeДокумент5 страниц2018overnutrition Ectopic Lipid and The Metabolic SyndromeLuminita HutanuОценок пока нет
- Obesity, Inflammation, and Atherosclerosis: Viviane Z. Rocha and Peter LibbyДокумент11 страницObesity, Inflammation, and Atherosclerosis: Viviane Z. Rocha and Peter LibbyFco Javier Sanchez PeñaОценок пока нет
- De Novo Lipogenesis As A Source of Second Messengers in Adipocytes (Hsiao y Guertin, 2019)Документ13 страницDe Novo Lipogenesis As A Source of Second Messengers in Adipocytes (Hsiao y Guertin, 2019)Andrea MoralesОценок пока нет
- Mammi 2012 YAAAДокумент8 страницMammi 2012 YAAAXochitemiqui YoacihualtОценок пока нет
- Importance of The Microenvironment and Mechanosensing in Adipose Tissue Biology 2022 ReviewДокумент29 страницImportance of The Microenvironment and Mechanosensing in Adipose Tissue Biology 2022 Review畏Оценок пока нет
- Chronic Lipo FatДокумент12 страницChronic Lipo FatDenisse ZayasОценок пока нет
- Makrofag Pada Jaringan Adiposa Obes Sebagai Penanda Terjadinya Resistensi InsulinДокумент6 страницMakrofag Pada Jaringan Adiposa Obes Sebagai Penanda Terjadinya Resistensi InsulinOris WicaksonoОценок пока нет
- Targeting Senescent Cells Enhances Adipogenesis and Metabolic Function in Old AgeДокумент19 страницTargeting Senescent Cells Enhances Adipogenesis and Metabolic Function in Old AgeirmaОценок пока нет
- Scheja & Heeren 2019 The Endocrine Function of Adipose Tissues in Health and Cardiometabolic DiseaseДокумент18 страницScheja & Heeren 2019 The Endocrine Function of Adipose Tissues in Health and Cardiometabolic DiseasevibhutiОценок пока нет
- Obesity, Adiposity, DyslipidemiaДокумент80 страницObesity, Adiposity, DyslipidemiaYehezkiel Dwi Putra100% (1)
- Impaired Protein Metabolism: Interlinks Between Obesity, Insulin Resistance and InflammationДокумент7 страницImpaired Protein Metabolism: Interlinks Between Obesity, Insulin Resistance and InflammationThamyres CristhinaОценок пока нет
- Adipose Organ Development and Remodeling (Cinti, 2018)Документ75 страницAdipose Organ Development and Remodeling (Cinti, 2018)VELEKAОценок пока нет
- Mekanisme Terjadinya Inflamasi Dan Stres Oksidatif Pada Obesitas Ana RahmawatiДокумент8 страницMekanisme Terjadinya Inflamasi Dan Stres Oksidatif Pada Obesitas Ana RahmawatimerkuriОценок пока нет
- Mental Consequences of ObesityДокумент7 страницMental Consequences of Obesitykeshav singhОценок пока нет
- Regulation of Stem Cell Differentiation in Adipose Tissue by Chronic InflammationДокумент13 страницRegulation of Stem Cell Differentiation in Adipose Tissue by Chronic InflammationSindy Noreima Nino VegaОценок пока нет
- Mediterranean - Diet - Endothelial - Function - and - Vascular - Inflammatory - Markers 2018 PDFДокумент4 страницыMediterranean - Diet - Endothelial - Function - and - Vascular - Inflammatory - Markers 2018 PDFPaulo CardosoОценок пока нет
- Artículos Originales: Fisiopatología de La Obesidad: Perspectiva Actual Pathophysiology of Obesity: Current ViewДокумент8 страницArtículos Originales: Fisiopatología de La Obesidad: Perspectiva Actual Pathophysiology of Obesity: Current ViewJosé Maturana SilvОценок пока нет
- Artículos Originales: Fisiopatología de La Obesidad: Perspectiva Actual Pathophysiology of Obesity: Current ViewДокумент8 страницArtículos Originales: Fisiopatología de La Obesidad: Perspectiva Actual Pathophysiology of Obesity: Current ViewDaniel SanchezОценок пока нет
- Fisiopatología de La Obesidad. Perspectiva ActualДокумент8 страницFisiopatología de La Obesidad. Perspectiva ActualMarcia Alvarez ZeballosОценок пока нет
- Obesity Is Associated With Macrophage Accumulation in Adipose TissueДокумент13 страницObesity Is Associated With Macrophage Accumulation in Adipose TissueRachel Lalaine Marie SialanaОценок пока нет
- Adipose Tissue Dysfunction As Determinant of Obesity-Associated Metabolic ComplicationsДокумент23 страницыAdipose Tissue Dysfunction As Determinant of Obesity-Associated Metabolic ComplicationsAli.m DaoodОценок пока нет
- Insight Into The Development of Obesity: Functional Alterations of Adipose-Derived Mesenchymal Stem CellsДокумент17 страницInsight Into The Development of Obesity: Functional Alterations of Adipose-Derived Mesenchymal Stem CellsJulia SCОценок пока нет
- Full Length Research Article: ISSN: 2230-9926Документ9 страницFull Length Research Article: ISSN: 2230-9926Somayeh Zaminpira100% (1)
- Nihms 599061Документ31 страницаNihms 599061herryОценок пока нет
- Muscles, Exercise and ObesityДокумент9 страницMuscles, Exercise and ObesityJohana FuentesОценок пока нет
- Artículo 2Документ15 страницArtículo 2Javier PereiraОценок пока нет
- Dynamics of Adipose Tissue Turnover in Human Metabolic Health and Disease (White Et Al., 2018)Документ7 страницDynamics of Adipose Tissue Turnover in Human Metabolic Health and Disease (White Et Al., 2018)Maria Luisa Sibajas ÁvilaОценок пока нет
- Increasing Muscle Mass To CДокумент8 страницIncreasing Muscle Mass To CCarlos MosqueraОценок пока нет
- Adipose Tissue, Inlammation (Meta-Inlammation) and Obesity ManagementДокумент18 страницAdipose Tissue, Inlammation (Meta-Inlammation) and Obesity ManagementHelena FabianiОценок пока нет
- Adiposa 2Документ27 страницAdiposa 2Nindya Agustin RОценок пока нет
- The Action of Vitamin D in Adipose TissueДокумент35 страницThe Action of Vitamin D in Adipose TissueMatheus MaltaОценок пока нет
- Laforest 2015 Adipocyte Size As A Determinant of Ms and Adipose Tissue DysfunctionДокумент13 страницLaforest 2015 Adipocyte Size As A Determinant of Ms and Adipose Tissue DysfunctionPaul SimononОценок пока нет
- Arsenic-Stimulated Lipolysis and Adipose Remodeling Is Mediated by G-Protein-Coupled ReceptorsДокумент10 страницArsenic-Stimulated Lipolysis and Adipose Remodeling Is Mediated by G-Protein-Coupled Receptorsevanconstantine77Оценок пока нет
- Lipid Accumulation in Non-Adipose Tissue and Lipotoxicity: N.A. Van Herpen, V.B. Schrauwen-HinderlingДокумент11 страницLipid Accumulation in Non-Adipose Tissue and Lipotoxicity: N.A. Van Herpen, V.B. Schrauwen-HinderlingEvan Permana PutraОценок пока нет
- HHS Public Access: Exercise and Metabolic Health: Beyond Skeletal MuscleДокумент21 страницаHHS Public Access: Exercise and Metabolic Health: Beyond Skeletal MuscleYaasar NazarОценок пока нет
- Obesity and Immune FunctionVF 19 - 20Документ53 страницыObesity and Immune FunctionVF 19 - 20bmvdgxjwh7Оценок пока нет
- Journal of Diabetes - 2019 - BeverlyДокумент3 страницыJournal of Diabetes - 2019 - BeverlyHeru SetiawanОценок пока нет
- Skin With MtabolicДокумент13 страницSkin With MtabolicRisky PerdanaОценок пока нет
- Adipogenesis A Necessary But Harmful StrategyДокумент27 страницAdipogenesis A Necessary But Harmful Strategydenny070111209Оценок пока нет
- SB7.1p HormonesДокумент9 страницSB7.1p HormonesHisokagenОценок пока нет
- Clinical Practice Guidelines: Biliary Cancer: ESMO Clinical Practice Guidelines For Diagnosis, Treatment and Follow-UpДокумент10 страницClinical Practice Guidelines: Biliary Cancer: ESMO Clinical Practice Guidelines For Diagnosis, Treatment and Follow-UpPutri MauraОценок пока нет
- The Allergic Response: Kathleen Nicole B. Bidua First Year ResidentДокумент44 страницыThe Allergic Response: Kathleen Nicole B. Bidua First Year ResidentNicole BiduaОценок пока нет
- A.1.1 Embriologi OtakДокумент28 страницA.1.1 Embriologi OtakoliviaОценок пока нет
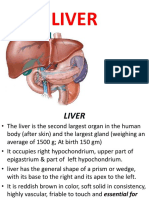
- LIVERДокумент18 страницLIVERShivaniLeela100% (3)
- Total Laparoscopic Hysterectomy (TLH) : Patient Information Patient InformationДокумент7 страницTotal Laparoscopic Hysterectomy (TLH) : Patient Information Patient InformationMUKUL SAXENAОценок пока нет
- Haemostasis: Case StudyДокумент19 страницHaemostasis: Case StudyTusabe FredОценок пока нет
- Human Anatomy 5th Edition Saladin Test BankДокумент47 страницHuman Anatomy 5th Edition Saladin Test Bankbevisnhan89cq0m100% (30)
- Anterior Triangle of NeckДокумент13 страницAnterior Triangle of NeckÑäd ÉèmОценок пока нет
- Notes On Antibodies PropertiesДокумент3 страницыNotes On Antibodies PropertiesBidur Acharya100% (1)
- LESSON EXEMPLAR IN SCIENCE 10. Co2Документ15 страницLESSON EXEMPLAR IN SCIENCE 10. Co2Hazel HazeОценок пока нет
- Pridevi PREPravkeДокумент8 страницPridevi PREPravkeLukaОценок пока нет
- Author 'S Accepted ManuscriptДокумент46 страницAuthor 'S Accepted ManuscriptRaul DoctoОценок пока нет
- Abo Blood Group SystemДокумент17 страницAbo Blood Group SystemCSC LanjipadarОценок пока нет
- Maslow and Collinge - Course of The Femoral Artery in The Mid - and Distal Thigh and Implications For Medial Approaches To The Distal Femur (2019)Документ5 страницMaslow and Collinge - Course of The Femoral Artery in The Mid - and Distal Thigh and Implications For Medial Approaches To The Distal Femur (2019)Kristijonas PetkevičiusОценок пока нет
- PL001-A1 Advance Steel PDFДокумент1 страницаPL001-A1 Advance Steel PDFpaúl quicañoОценок пока нет
- Muscle TissueДокумент20 страницMuscle TissueDheeraj MenezesОценок пока нет
- The Paris System For Reporting Urinary CytologyДокумент66 страницThe Paris System For Reporting Urinary CytologyRawa MuhsinОценок пока нет
- Special Senses) 3. Photoreceptors - Rods and Cones - KeyДокумент1 страницаSpecial Senses) 3. Photoreceptors - Rods and Cones - Keyranag59100Оценок пока нет
- Surgical Anatomy of The Biliary TractДокумент5 страницSurgical Anatomy of The Biliary TracthoangducnamОценок пока нет
- PBSДокумент72 страницыPBSKavi KaviОценок пока нет
- Dr. Witra Irfan, SP.B (K) V - DVTДокумент29 страницDr. Witra Irfan, SP.B (K) V - DVTmuhammad azharanОценок пока нет
- Cranial Nerve: Dr. Dian Prasetyo Wibisono, M.SCДокумент49 страницCranial Nerve: Dr. Dian Prasetyo Wibisono, M.SCgeneОценок пока нет
- Chapter 1 Anaphy MariebДокумент26 страницChapter 1 Anaphy MariebKeanna ZurriagaОценок пока нет
- Ultrasound Measurements Data Communication: Aloka Co., LTDДокумент8 страницUltrasound Measurements Data Communication: Aloka Co., LTDRama Tenis CopecОценок пока нет
- Lesson 1: The Curriculum Guide ObjectivesДокумент48 страницLesson 1: The Curriculum Guide ObjectivesGodwin Jerome ReyesОценок пока нет
- Respiratory and Circulatory System Detailed Lesson PlanДокумент23 страницыRespiratory and Circulatory System Detailed Lesson PlanEdnalyn LicotОценок пока нет
- Head To Toe AssessmentДокумент3 страницыHead To Toe AssessmentKimberly Witt WhitlockОценок пока нет
- Govt. Girls High School Pura Heeran, SialkotДокумент7 страницGovt. Girls High School Pura Heeran, SialkotFizza cooks foodОценок пока нет
- 11th (D) Test 2Документ2 страницы11th (D) Test 2Deeyansh DahatОценок пока нет