Вам также может понравиться
- Factores AbióticosДокумент7 страницFactores AbióticosIsaac Herbas MattosОценок пока нет
- LAB Morfología de Hongos y BacteriasДокумент16 страницLAB Morfología de Hongos y BacteriasWilliam Reyes PeñaОценок пока нет
- Camilo Leal Ecosistemas Lénticos y Lóticos 2 PDFДокумент5 страницCamilo Leal Ecosistemas Lénticos y Lóticos 2 PDFCamilo Leal100% (1)
- 001 Fisica Informe de Caja de ResonanciaДокумент6 страниц001 Fisica Informe de Caja de ResonanciaYolimarVelezОценок пока нет
- Tubo de Kundt PDFДокумент12 страницTubo de Kundt PDFMarisol Ortiz GОценок пока нет
- Responsabilidad Del Educador SujetoДокумент4 страницыResponsabilidad Del Educador SujetoPamela GarciaОценок пока нет
- Planteamiento Del ProblemaДокумент6 страницPlanteamiento Del ProblemaLuis OdicioОценок пока нет
- Maloka PDFДокумент15 страницMaloka PDFjavier bОценок пока нет
- Trabajo Final InvestigativoДокумент44 страницыTrabajo Final Investigativoorleidys gomez caicedoОценок пока нет
- Maloka ElectromagnetismoДокумент10 страницMaloka ElectromagnetismoSergio Daniel Rodriguez Preciado0% (1)
- Trabajo Practico Péndulo SimpleДокумент4 страницыTrabajo Practico Péndulo SimpleLeonardo A. FunesОценок пока нет
- Una Visión Holistica de La Investigación PedagógicaДокумент4 страницыUna Visión Holistica de La Investigación PedagógicaMauro MorenoОценок пока нет
- EJEMPLO-diapositivas Cuasi ExperimentalДокумент5 страницEJEMPLO-diapositivas Cuasi ExperimentalHebert MamaniОценок пока нет
- Programa NanomaterialesДокумент8 страницPrograma NanomaterialesMaxi GonzalezОценок пока нет
- Taller Moléculas y Seres Vivos Juana FДокумент5 страницTaller Moléculas y Seres Vivos Juana FJUANA VALENTINA FRANCO CORREDORОценок пока нет
- Introducción de La MitosisДокумент5 страницIntroducción de La MitosisSneyder Leyton Cárdenas100% (1)
- Philippe Meirieu Transformacion PedagogicaДокумент12 страницPhilippe Meirieu Transformacion Pedagogicajairo alberto galindo cuestaОценок пока нет
- Informe Del OjoДокумент4 страницыInforme Del OjoJonathan Enríquez0% (1)
- ProtoplasmaДокумент9 страницProtoplasmaChristopher Guevara CheОценок пока нет
- Propiedades de Las Células CancerigenasДокумент9 страницPropiedades de Las Células CancerigenasfausОценок пока нет
- Indices de Diversidad EcosistemicaДокумент2 страницыIndices de Diversidad EcosistemicaJuan Carlos Giraldo RiosОценок пока нет
- Practica 1 Fuerza ElectricaДокумент11 страницPractica 1 Fuerza ElectricaCessar Augusto Vergara ArdilaОценок пока нет
- Ondas Estacionarias en Tubos AbiertosДокумент7 страницOndas Estacionarias en Tubos AbiertosFrancy Cristina Ramirez BedoyaОценок пока нет
- Demostracion de Las Leyes de A TermodinamicaДокумент20 страницDemostracion de Las Leyes de A TermodinamicaJoel Quisuruco CasasОценок пока нет
- 8a-Campos Magneticos Estaticos - HДокумент25 страниц8a-Campos Magneticos Estaticos - HMario Javier PradaОценок пока нет
- Manual PlactonДокумент60 страницManual PlactonNathaliaZarzaОценок пока нет
- Laboratorio Morfologia Del OjoДокумент14 страницLaboratorio Morfologia Del OjoNiceth Natalia Ricaurte MorenoОценок пока нет
- Evolucion de La MitosisДокумент5 страницEvolucion de La MitosisLina Munoz100% (1)
- Informe Ondas Sonoras PDFДокумент17 страницInforme Ondas Sonoras PDFJimmy Alexander Ochoa RuizОценок пока нет
- Fibra OpticaДокумент8 страницFibra OpticaSergio BoccheciampeОценок пока нет
- Practica de Laboratorio # 4-Estudio de Las Lentes-2012505Документ5 страницPractica de Laboratorio # 4-Estudio de Las Lentes-2012505Ruben100% (1)
- Impulso NerviosoДокумент10 страницImpulso NerviosomimiОценок пока нет
- Estructura Interna de La MateriaДокумент4 страницыEstructura Interna de La MateriaLuis Carlos MosqueraОценок пока нет
- Guia Basica de Quimica 4Документ3 страницыGuia Basica de Quimica 4ovidio martinez100% (1)
- Informe de Extracción de Adn BiologíaДокумент5 страницInforme de Extracción de Adn BiologíaCarito Lisseth BorjaОценок пока нет
- Transporte en La MembranaДокумент10 страницTransporte en La MembranaCRISTIAN MANUEL SAAVEDRA RODRIGUEZОценок пока нет
- Modelos de ProduccionДокумент6 страницModelos de Produccionhector cedeñoОценок пока нет
- Prac 6 Permeabilidad Celular 1Документ18 страницPrac 6 Permeabilidad Celular 1freshaОценок пока нет
- Tema 3. Señalización Celular PDFДокумент18 страницTema 3. Señalización Celular PDFAelita RuseelОценок пока нет
- Microscopia de Contraste de FasesДокумент7 страницMicroscopia de Contraste de FasesIvan Salas Gaeta100% (1)
- Practica OsmosisДокумент6 страницPractica OsmosisJhonatan Santos MartínezОценок пока нет
- Fenómenos de MembranasДокумент8 страницFenómenos de MembranasArlex Lara Sánchez100% (1)
- Articulo Cientifico Reconocimiento y Clasificacion de SuelosДокумент14 страницArticulo Cientifico Reconocimiento y Clasificacion de SuelosAnyelii08Оценок пока нет
- Secuencia 2° Digestivo Uruguay EducaДокумент9 страницSecuencia 2° Digestivo Uruguay EducaIanara RodríguezОценок пока нет
- La MitocondriaДокумент4 страницыLa MitocondriaDavid GatoОценок пока нет
- Laboratorio06 Osmosis en Plantas y AnimalesДокумент6 страницLaboratorio06 Osmosis en Plantas y AnimalesSalomón Marín GarnicaОценок пока нет
- Informe MicrobiologíaДокумент5 страницInforme MicrobiologíaLiizhiiz Vaca100% (1)
- Sistema OsteoartromuscularДокумент5 страницSistema OsteoartromuscularAdriánGonzálezОценок пока нет
- Morbilidad Infantil PDFДокумент27 страницMorbilidad Infantil PDFRebeca ChacínОценок пока нет
- Teorías Sobre El Origen de La Célula EucariotaДокумент1 страницаTeorías Sobre El Origen de La Célula EucariotaMauro Villarrubia67% (3)
- Encéfalo: Prosencéfalo, Mesencéfalo, RombencéfaloДокумент13 страницEncéfalo: Prosencéfalo, Mesencéfalo, RombencéfaloSthefanny MedinaОценок пока нет
- Informe de Laboratorio de Biología - MicroscopíaДокумент11 страницInforme de Laboratorio de Biología - MicroscopíameromachofullОценок пока нет
- El Sistema o Aparato Excretor de Los AnimalesДокумент5 страницEl Sistema o Aparato Excretor de Los AnimalesEvelynОценок пока нет
- Los Cinco ReinosДокумент12 страницLos Cinco ReinosJhoselyn Esther GarcíaОценок пока нет
- Quiralidad en La Química La Física y La Biología: by Jean Paul Tunjano CallejasДокумент8 страницQuiralidad en La Química La Física y La Biología: by Jean Paul Tunjano CallejasJean Paul HessОценок пока нет
- Anelidos Informe Nota 95Документ8 страницAnelidos Informe Nota 95Noemi DuranОценок пока нет
- Transporte A Través de La Membrana Celular o PlasmáticaДокумент6 страницTransporte A Través de La Membrana Celular o PlasmáticaFco ZalgadoОценок пока нет
- Superficies EquipotencialesДокумент10 страницSuperficies EquipotencialesGabriel PintoОценок пока нет
- Celula HipertonicaДокумент33 страницыCelula HipertonicaErika Viviana MoncadaОценок пока нет
- Contraccion Muscular ExpoДокумент20 страницContraccion Muscular ExpoElisabet KentОценок пока нет
- EdulcorantesДокумент5 страницEdulcorantesviaereaОценок пока нет
- Seminario Integracion MetabolicaДокумент23 страницыSeminario Integracion MetabolicafawingpeОценок пока нет
- Fibra DieteticaДокумент12 страницFibra DieteticaIvette DomínguezОценок пока нет
- Antropometria Universitarios PDFДокумент6 страницAntropometria Universitarios PDFRoberto Esparza SolisОценок пока нет
- Perspectiva y Patrónde AlimentaciДокумент5 страницPerspectiva y Patrónde AlimentaciRoberto Esparza SolisОценок пока нет
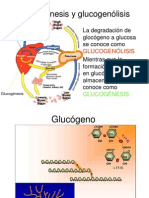
- 11GLUCOGENOLISISДокумент30 страниц11GLUCOGENOLISISRoberto Esparza Solis100% (1)
- EjerciciosДокумент3 страницыEjerciciosJazz Cube100% (2)
- Informe Proyecto Muro de ContencionДокумент23 страницыInforme Proyecto Muro de ContencionjessОценок пока нет
- CB33-37 Química Orgánica KjaskxДокумент12 страницCB33-37 Química Orgánica Kjaskxjorge0% (1)
- Proyecto Rehabilitacion VialДокумент92 страницыProyecto Rehabilitacion VialMAIRA ORTIZОценок пока нет
- Lectura Comprension CTДокумент2 страницыLectura Comprension CTMoroni EspinosaОценок пока нет
- Los Diagramas de Tubería e Instrumentación MДокумент15 страницLos Diagramas de Tubería e Instrumentación MMinerva Torres100% (2)
- Minicargadores 236 DДокумент20 страницMinicargadores 236 DdianaОценок пока нет
- Cálculo de Pérdidas de CargaДокумент2 страницыCálculo de Pérdidas de CargaAlexander Molina YateОценок пока нет
- Prueba T de StudentДокумент7 страницPrueba T de StudentDavid Palacios IngaОценок пока нет
- Absorción Partículas BetaДокумент4 страницыAbsorción Partículas Betaalejandro amadoОценок пока нет
- Cap 1Документ8 страницCap 1fenixseravgonОценок пока нет
- Clase 1Документ7 страницClase 1Afro SamuraiОценок пока нет
- Analisis-Granulometrico-Fundamentos - (1) (1) (Autoguardado)Документ15 страницAnalisis-Granulometrico-Fundamentos - (1) (1) (Autoguardado)Cordova RaphaelОценок пока нет
- Problemas de Práctica #1Документ7 страницProblemas de Práctica #1Daniel CandanedoОценок пока нет
- AlgebraДокумент10 страницAlgebraCésar GarcíaОценок пока нет
- Practica de Laboratorio 2Документ9 страницPractica de Laboratorio 2Edgar Morales SilverioОценок пока нет
- Diseño en Compresión PDFДокумент64 страницыDiseño en Compresión PDFJuniorLeonОценок пока нет
- Angulo de Elevación y DepresiónДокумент2 страницыAngulo de Elevación y DepresiónMiguel Inti MorenoОценок пока нет
- Procedimiento Programa de Calibración de Equipos de MediciónДокумент4 страницыProcedimiento Programa de Calibración de Equipos de Mediciónlehoaqp100% (2)
- Cromado Industrial PDFДокумент4 страницыCromado Industrial PDFOswaldo GuerraОценок пока нет
- Homew3 PDFДокумент3 страницыHomew3 PDFDavid LabradorОценок пока нет
- Actividades Colaborativo Capitulo 2 - Teoria de Numeros 2017-1Документ6 страницActividades Colaborativo Capitulo 2 - Teoria de Numeros 2017-1Jonathan Ardila VergaraОценок пока нет
- Fruta ConfitadaДокумент8 страницFruta ConfitadaFrancisco Luicho LeonОценок пока нет
- Programacion Metodo Del Lumen Version 2Документ6 страницProgramacion Metodo Del Lumen Version 2Adrian QuintanillaОценок пока нет
- Curvas Voltaje CorrienteДокумент46 страницCurvas Voltaje CorrienteGabrielaAlatristaОценок пока нет
- Agua Caliente SanitariaДокумент15 страницAgua Caliente SanitariaJose Juarez AlquizarОценок пока нет
- Reacciones QuímicasДокумент12 страницReacciones QuímicasMariel BazánОценок пока нет
- Arrancador Suave y Variador de VelocidadДокумент5 страницArrancador Suave y Variador de VelocidadcrapnawОценок пока нет
- Cinemática I PDFДокумент8 страницCinemática I PDFValeria Gutierrez OrellanaОценок пока нет