Вам также может понравиться
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeОт EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeРейтинг: 4 из 5 звезд4/5 (5794)
- Delirium IncessabilisДокумент24 страницыDelirium IncessabilisMiguel ClaramuntОценок пока нет
- The Little Book of Hygge: Danish Secrets to Happy LivingОт EverandThe Little Book of Hygge: Danish Secrets to Happy LivingРейтинг: 3.5 из 5 звезд3.5/5 (400)
- A Rose Midst Rubble PDFДокумент29 страницA Rose Midst Rubble PDFMiguel ClaramuntОценок пока нет
- A Rose Midst Rubble PDFДокумент29 страницA Rose Midst Rubble PDFMiguel ClaramuntОценок пока нет
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceОт EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceРейтинг: 4 из 5 звезд4/5 (895)
- Acanthaceae of SonoraДокумент116 страницAcanthaceae of SonoraMiguel ClaramuntОценок пока нет
- The Yellow House: A Memoir (2019 National Book Award Winner)От EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Рейтинг: 4 из 5 звезд4/5 (98)
- Coleman 2010 - Delta For BeginnersДокумент75 страницColeman 2010 - Delta For BeginnersMiguel ClaramuntОценок пока нет
- The Emperor of All Maladies: A Biography of CancerОт EverandThe Emperor of All Maladies: A Biography of CancerРейтинг: 4.5 из 5 звезд4.5/5 (271)
- Week 8 - Leaves and StemsДокумент77 страницWeek 8 - Leaves and StemsPrincess De LeonОценок пока нет
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryОт EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryРейтинг: 3.5 из 5 звезд3.5/5 (231)
- MansdrfДокумент2 страницыMansdrfmansi bavliyaОценок пока нет
- Never Split the Difference: Negotiating As If Your Life Depended On ItОт EverandNever Split the Difference: Negotiating As If Your Life Depended On ItРейтинг: 4.5 из 5 звезд4.5/5 (838)
- Powder Microscopy of Crude Drugs PDFДокумент52 страницыPowder Microscopy of Crude Drugs PDFSunitha Katta100% (5)
- आकाश्वल्ली या अमरबेलДокумент10 страницआकाश्वल्ली या अमरबेलaadityaОценок пока нет
- Giant Sequoia, Sequoiadendron Giganteum: Where Do Trees Get Their Mass From?Документ19 страницGiant Sequoia, Sequoiadendron Giganteum: Where Do Trees Get Their Mass From?travis strauseОценок пока нет
- Botany 101Документ7 страницBotany 101Gladdin ChloeОценок пока нет
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureОт EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureРейтинг: 4.5 из 5 звезд4.5/5 (474)
- Instant Download Test Bank For Voyages in World History Volume 2 3rd Edition by Hansen PDF ScribdДокумент32 страницыInstant Download Test Bank For Voyages in World History Volume 2 3rd Edition by Hansen PDF ScribdDaniel Johnson100% (12)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaОт EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaРейтинг: 4.5 из 5 звезд4.5/5 (266)
- Botany Practice Test I - Answer KeyДокумент4 страницыBotany Practice Test I - Answer KeyIrene Miranda FloresОценок пока нет
- The Unwinding: An Inner History of the New AmericaОт EverandThe Unwinding: An Inner History of the New AmericaРейтинг: 4 из 5 звезд4/5 (45)
- Tudung AkarДокумент17 страницTudung AkarSyifa SariОценок пока нет
- LG 2.4 - Selected Plant Diseases (Part 2) PDFДокумент52 страницыLG 2.4 - Selected Plant Diseases (Part 2) PDFIzaiah Ishe CastroОценок пока нет
- Team of Rivals: The Political Genius of Abraham LincolnОт EverandTeam of Rivals: The Political Genius of Abraham LincolnРейтинг: 4.5 из 5 звезд4.5/5 (234)
- 11th Botany NotesДокумент43 страницы11th Botany Notesruben psОценок пока нет
- XII - CBSE - Worksheet - Anatomy of Flowering Plants - Answer KeyДокумент1 страницаXII - CBSE - Worksheet - Anatomy of Flowering Plants - Answer KeyKishor KumarОценок пока нет
- Root Modifications Storage RootsДокумент17 страницRoot Modifications Storage Rootsmbg1110100% (1)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyОт EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyРейтинг: 3.5 из 5 звезд3.5/5 (2259)
- Somatic Structures of FungiДокумент2 страницыSomatic Structures of FungiSajjad Hossain Shuvo100% (1)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreОт EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreРейтинг: 4 из 5 звезд4/5 (1090)
- Digital Atlas of Woody Plants, For Web Site, 27 Apr 2020Документ736 страницDigital Atlas of Woody Plants, For Web Site, 27 Apr 2020LauraОценок пока нет
- Lower Higher Plants TaxonomyДокумент36 страницLower Higher Plants TaxonomyIvka Zemiakova100% (2)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersОт EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersРейтинг: 4.5 из 5 звезд4.5/5 (344)
- 23Документ3 страницы23gem68megОценок пока нет
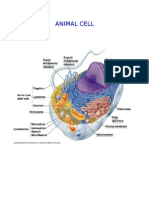
- Animal CellДокумент5 страницAnimal CellNan Fen Jin Sabolarce100% (1)
- CELERY LAB - Structure and Function of A PlantДокумент5 страницCELERY LAB - Structure and Function of A PlantShyla Fave EnguitoОценок пока нет
- Form 5 Biology - Online Revision Quiz #4 - Reproduction in Plants - PFLДокумент4 страницыForm 5 Biology - Online Revision Quiz #4 - Reproduction in Plants - PFLParco ChanОценок пока нет
- (I) Short-Day Plants (SDP)Документ6 страниц(I) Short-Day Plants (SDP)keerthi vasanОценок пока нет
- Lec19 PDFДокумент11 страницLec19 PDFsyakiraОценок пока нет
- Edu246 Botany 2Документ8 страницEdu246 Botany 2ANNE MARGUERITE BARETEОценок пока нет
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)От EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Рейтинг: 4.5 из 5 звезд4.5/5 (121)
- Andreaeales (Lantern Moss)Документ12 страницAndreaeales (Lantern Moss)greeshma vasu100% (1)
- BOT 121 TranspirationДокумент7 страницBOT 121 TranspirationSakshi SargarОценок пока нет
- Biology The Dynamic Science 4th Edition Russell Solutions Manual 1Документ10 страницBiology The Dynamic Science 4th Edition Russell Solutions Manual 1matthew100% (30)
- Poster Klasifikasi Filogeni LlumutДокумент1 страницаPoster Klasifikasi Filogeni LlumutsuhendarsomawijayaОценок пока нет
- Unnathi-Biology Inner Pages 8-7-23 FinalДокумент188 страницUnnathi-Biology Inner Pages 8-7-23 Finalramjaan5000Оценок пока нет
- Chapter 1Документ21 страницаChapter 1chapter 1Оценок пока нет
- Secondary Growth in Dicot Stem (With Diagram) : NavigationДокумент41 страницаSecondary Growth in Dicot Stem (With Diagram) : NavigationNasser Manungka SabandalОценок пока нет