Вам также может понравиться
- Curso Panaderia Basico RotaryДокумент9 страницCurso Panaderia Basico RotaryFernanda ValentinaОценок пока нет
- Flujo Monofásico en Sistemas Homogéneos Lineales y RadialesДокумент8 страницFlujo Monofásico en Sistemas Homogéneos Lineales y RadialesAlessandro GarciaОценок пока нет
- P6. Reactivo Limitante-Converted ReparadoДокумент5 страницP6. Reactivo Limitante-Converted ReparadoChristopher MorenoОценок пока нет
- Inflamación y Reparación - Reparación de TejidosДокумент10 страницInflamación y Reparación - Reparación de TejidosEmmanuelОценок пока нет
- Cuadro de Medios de CultivoДокумент5 страницCuadro de Medios de CultivoVanesa Cx SlumberОценок пока нет
- Tinción de Flagelos (Paco)Документ13 страницTinción de Flagelos (Paco)Marco OlguínОценок пока нет
- Analisis Articulos Ley Organica de Los Procesos AdministrativosДокумент14 страницAnalisis Articulos Ley Organica de Los Procesos AdministrativosChavela DepablosОценок пока нет
- Embarazos en Edades ExtremasДокумент17 страницEmbarazos en Edades ExtremasChavela DepablosОценок пока нет
- Agar Sangre Base Infusiòn ValtekДокумент2 страницыAgar Sangre Base Infusiòn ValtekAnderson Obed Alvarez MondragonОценок пока нет
- Estruct Bacteria Apuntes PDFДокумент10 страницEstruct Bacteria Apuntes PDFBioloscarОценок пока нет
- Estilosde Investigacionsobrelaclaseobrera Dela GarzaДокумент18 страницEstilosde Investigacionsobrelaclaseobrera Dela GarzaChavela DepablosОценок пока нет
- Héroes de VenezuelaДокумент3 страницыHéroes de VenezuelaChavela Depablos100% (2)
- Tradiciones de Año Viejo y Año Nuevo en VenezuelaДокумент1 страницаTradiciones de Año Viejo y Año Nuevo en VenezuelaChavela DepablosОценок пока нет
- Archivetemp6. EVALUANDO EL APRENDIZAJEДокумент2 страницыArchivetemp6. EVALUANDO EL APRENDIZAJEIngrid Gisela TUMBACO TAUTASОценок пока нет
- Formulacion de AgaresДокумент12 страницFormulacion de AgaresAntonello LedezmaОценок пока нет
- Invernaderos MeridaДокумент10 страницInvernaderos MeridaJosé Luis RomeroОценок пока нет
- Práctica Química 2008-IIДокумент58 страницPráctica Química 2008-IILiaLeonardoОценок пока нет
- Ensayo de TraccionДокумент14 страницEnsayo de TraccionMauricio AguirreОценок пока нет
- Operaciones de ReacciónДокумент28 страницOperaciones de ReacciónMarcioLazОценок пока нет
- Proteccion Catodica (Monografia) - CorrosionДокумент36 страницProteccion Catodica (Monografia) - CorrosionBenjamin Palma Inocente100% (1)
- Capítulo 15A - Grupo 3 - Ciencia de Los MaterialesДокумент2 страницыCapítulo 15A - Grupo 3 - Ciencia de Los MaterialesRolexОценок пока нет
- Ugt Diseño TronadasДокумент5 страницUgt Diseño TronadasVictor HernanОценок пока нет
- Proceso de liofilización de caféДокумент3 страницыProceso de liofilización de caféMartha Isabel Vivas SandovalОценок пока нет
- 1 Manual de OrganizacionДокумент28 страниц1 Manual de OrganizacionSheyla Yesenia Garay ValverdeОценок пока нет
- Exposición - LavanderiaДокумент4 страницыExposición - LavanderiaKatherin SalasОценок пока нет
- Natal Branco PARTITURA 3Документ4 страницыNatal Branco PARTITURA 3Cadu BarcelosОценок пока нет
- Investigacion Centrales TermicasДокумент7 страницInvestigacion Centrales TermicasSamuel DiazОценок пока нет
- Metformina d2Документ7 страницMetformina d2Luis Guillermo Sanchez LezamaОценок пока нет
- Pulsos LuminicosДокумент22 страницыPulsos LuminicosAndrea Reyes Tello100% (1)
- Diseño de mezclas asfálticas método Marshall: procedimiento, cálculos e interpretación de resultadosДокумент10 страницDiseño de mezclas asfálticas método Marshall: procedimiento, cálculos e interpretación de resultadosgabymorga7Оценок пока нет
- EFECTO INVERNADERO Y CALENTAMIENTO GLOBALДокумент17 страницEFECTO INVERNADERO Y CALENTAMIENTO GLOBALHugo LauОценок пока нет
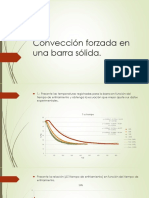
- Convección forzada en barra sólida: coeficiente de transferencia de energíaДокумент12 страницConvección forzada en barra sólida: coeficiente de transferencia de energíaKarliita LomОценок пока нет
- FORJAДокумент26 страницFORJAoscarhereОценок пока нет
- Cofactores Enzimáticos InorgánicosДокумент6 страницCofactores Enzimáticos InorgánicosRos HernandezОценок пока нет
- Guía prácticas biología molecularДокумент9 страницGuía prácticas biología molecularC_Dante33% (3)
- Preparación y Valoración de Nitrato de Plata 0Документ3 страницыPreparación y Valoración de Nitrato de Plata 0Edd de La ComarcaОценок пока нет
- INTA Pergamino Mezclas de Tanque y Prueba de CompatibilidadДокумент2 страницыINTA Pergamino Mezclas de Tanque y Prueba de CompatibilidadPedro Daniel LeivaОценок пока нет
- Ficha Tecnica Sulfato de SodioДокумент5 страницFicha Tecnica Sulfato de SodioRaúl Villagrán RozasОценок пока нет
- Piña La Piña Es Una Fruta Cítrica Llena de Vitaminas e Ideal para Una Dieta SaludableДокумент5 страницPiña La Piña Es Una Fruta Cítrica Llena de Vitaminas e Ideal para Una Dieta Saludablejessica roxannaОценок пока нет