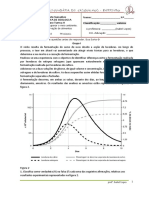
Вам также может понравиться
- Método de Escultura em Bloco de Cera-Incisivo Central Su Perior-21Документ15 страницMétodo de Escultura em Bloco de Cera-Incisivo Central Su Perior-21Cleisson Sampaio91% (11)
- CIPE Versão 1.0 PDFДокумент238 страницCIPE Versão 1.0 PDFMariana MargaridoОценок пока нет
- O Barroco em Portugal: Arte, Arquitetura e CaracterísticasДокумент5 страницO Barroco em Portugal: Arte, Arquitetura e CaracterísticasMariana Margarido100% (1)
- Ficha Informativa Sintese Proteica PDFДокумент6 страницFicha Informativa Sintese Proteica PDFcarlbisp1Оценок пока нет
- Preparação caseira de sabão a partir de gordurasДокумент4 страницыPreparação caseira de sabão a partir de gordurasMariana MargaridoОценок пока нет
- Relatório de QuímicaДокумент10 страницRelatório de QuímicaMariana MargaridoОценок пока нет
- Preparação caseira de sabão a partir de gordurasДокумент4 страницыPreparação caseira de sabão a partir de gordurasMariana MargaridoОценок пока нет
- 101 Técnicas para Ser Bom de CamaДокумент120 страниц101 Técnicas para Ser Bom de CamaEdson Bispo100% (3)
- Lista de Exercicios - BotanicaДокумент35 страницLista de Exercicios - BotanicaWilliasFariasОценок пока нет
- Controle de Qualidade em Urinálise PDFДокумент8 страницControle de Qualidade em Urinálise PDFJulio Cesar Schlemper60% (5)
- Escala de Dor LANNS E EVA PDFДокумент2 страницыEscala de Dor LANNS E EVA PDFHelena BritesОценок пока нет
- Ficha Informativa #1 Património GenéticoДокумент11 страницFicha Informativa #1 Património GenéticoPaulinhaa MesquitaОценок пока нет
- 06 Apostila Genetica IIДокумент5 страниц06 Apostila Genetica IIEduardo NascimentoОценок пока нет
- Património GenéticoДокумент7 страницPatrimónio GenéticoPaulo Silva CardosoОценок пока нет
- BIO12 Ficha - FINALДокумент8 страницBIO12 Ficha - FINALDulce SousaОценок пока нет
- Laboratório Sobre A Reprodução Sexuada Dos Seres Vivos - Reprodução Sexuada PDFДокумент38 страницLaboratório Sobre A Reprodução Sexuada Dos Seres Vivos - Reprodução Sexuada PDFNuno CâmaraОценок пока нет
- Teste 3Документ6 страницTeste 3zazasОценок пока нет
- 11-Hereditariedade ExercДокумент2 страницы11-Hereditariedade ExercMaria Inês BernardesОценок пока нет
- Ficha Formativa 3Документ5 страницFicha Formativa 3Ana BaptistaОценок пока нет
- Lista GeneticaДокумент5 страницLista GeneticaProfessora Carol100% (2)
- Biologia 12o - Trabalhos de MorganДокумент4 страницыBiologia 12o - Trabalhos de MorganJoana UrbanoОценок пока нет
- Avaliação de Biologia sobre GenéticaДокумент6 страницAvaliação de Biologia sobre GenéticaLara SantosОценок пока нет
- 2º Teste PDFДокумент7 страниц2º Teste PDFMaria MaiaОценок пока нет
- Teoria Cromossómica e Os Trabalhos de MorganДокумент39 страницTeoria Cromossómica e Os Trabalhos de MorganTricksterОценок пока нет
- FF6 - LetaisДокумент2 страницыFF6 - LetaisDavid MrspОценок пока нет
- Exercícios de Mutações e Técnicas de Engenharia GenéticaДокумент11 страницExercícios de Mutações e Técnicas de Engenharia GenéticaKarla KarrascoОценок пока нет
- DNA, genes e cromossomos: estrutura e hereditariedadeДокумент21 страницаDNA, genes e cromossomos: estrutura e hereditariedadePedro Vieira Martins CostaОценок пока нет
- Hereditariedade Biologia 12oДокумент4 страницыHereditariedade Biologia 12oteresa102100% (1)
- Ficha de Trabalho de Biologia 12º AnoДокумент7 страницFicha de Trabalho de Biologia 12º AnoAnonymous dPwFX8TIBMОценок пока нет
- Transmissão hereditária codominante visonsДокумент3 страницыTransmissão hereditária codominante visonsAna Catarina DiasОценок пока нет
- DownloadДокумент17 страницDownloadJoão PortelaОценок пока нет
- Ficha HereditariedadeДокумент5 страницFicha HereditariedadePaula FernandesОценок пока нет
- Exercícios GenéticaДокумент2 страницыExercícios GenéticaCharles Cabral25% (8)
- Ciclos de vida de organismosДокумент22 страницыCiclos de vida de organismosCarina GasparОценок пока нет
- DIVERSIDADE BIOLOGICA 10º AnoДокумент9 страницDIVERSIDADE BIOLOGICA 10º AnoMaria Candida SantosОценок пока нет
- Ficha de Trabalho - Unidade 2Документ4 страницыFicha de Trabalho - Unidade 2beatriz magalhaesОценок пока нет
- Sistema RhesusДокумент8 страницSistema RhesusblacksmokerОценок пока нет
- Reprodução sexuada e meiose em detalheДокумент28 страницReprodução sexuada e meiose em detalheBernardo MouratoОценок пока нет
- Segunda Lei de Mendel em questões de genéticaДокумент1 страницаSegunda Lei de Mendel em questões de genéticabarreiros8Оценок пока нет
- Análise da primeira lei de Mendel em um teste de BiologiaДокумент5 страницAnálise da primeira lei de Mendel em um teste de Biologiagelu1999Оценок пока нет
- 8-Ficha de Trabalho Ciclo SexualДокумент8 страниц8-Ficha de Trabalho Ciclo SexualNathaly AlvesОценок пока нет
- ESCOLA SECUNDÁRIA DE CASQUILHOS – BARREIRO: Fermentação de leveduras selvagens e de cultivoДокумент9 страницESCOLA SECUNDÁRIA DE CASQUILHOS – BARREIRO: Fermentação de leveduras selvagens e de cultivoExplicações Matemática Biologia FunchalОценок пока нет
- Estudo Drigido RESPOSTASДокумент18 страницEstudo Drigido RESPOSTASAstreОценок пока нет
- Kayla Santos - Genética I - Termologia e Primeira Lei de MendelДокумент6 страницKayla Santos - Genética I - Termologia e Primeira Lei de MendelKayla CpmОценок пока нет
- DNA e Síntese de Proteínas Biologia 11oДокумент34 страницыDNA e Síntese de Proteínas Biologia 11oqqqqwwwweeeerrrrtttt100% (1)
- Imunidade e Controlo de DoençasДокумент4 страницыImunidade e Controlo de DoençasDavid MrspОценок пока нет
- Avaliação Iii Unidade Vespertino BДокумент4 страницыAvaliação Iii Unidade Vespertino BRamalde SantanaОценок пока нет
- DNA e Síntese Proteica: Resumo da Biologia 11oДокумент25 страницDNA e Síntese Proteica: Resumo da Biologia 11ocarlbisp100% (1)
- FT4 - Sintese Proteíca:ciclo CelularДокумент18 страницFT4 - Sintese Proteíca:ciclo Celularjose eiraОценок пока нет
- Biologia 2Документ11 страницBiologia 2Cauan OkumuraОценок пока нет
- GenéticaДокумент7 страницGenéticaGabriel MirandaОценок пока нет
- Ficha Diagnóstico TAS BiologiaДокумент3 страницыFicha Diagnóstico TAS BiologiaRui NunesОценок пока нет
- Bio12º Ano - Manipulação FertilidadeДокумент4 страницыBio12º Ano - Manipulação FertilidadeSara RodriguesОценок пока нет
- Regulação e alteração do material genético na Biologia do 12o anoДокумент2 страницыRegulação e alteração do material genético na Biologia do 12o anoMaria Alcina BarataОценок пока нет
- Fichaformativa U2 TransmissãodecaraterísticashereditáriasДокумент3 страницыFichaformativa U2 Transmissãodecaraterísticashereditáriaslol lolОценок пока нет
- 10-Ciclos de VidasДокумент75 страниц10-Ciclos de VidasmanuelafonsecaОценок пока нет
- Ficha 2 HEREDITARIEDADE - CorreçãoДокумент5 страницFicha 2 HEREDITARIEDADE - CorreçãoCarlos LopesОценок пока нет
- DNA, RNA e Síntese ProteicaДокумент4 страницыDNA, RNA e Síntese ProteicaHelena Alves100% (1)
- Mitose e ciclo celularДокумент5 страницMitose e ciclo celularLara MesquitaОценок пока нет
- Genética de traços hereditários e sistemas sanguíneosДокумент8 страницGenética de traços hereditários e sistemas sanguíneospaulohnunes0% (1)
- Transmissão hereditáriaДокумент233 страницыTransmissão hereditáriaConceição PintoОценок пока нет
- Estrutura dos cromossomas eucarióticosДокумент14 страницEstrutura dos cromossomas eucarióticosinesОценок пока нет
- Variações Das Leis de Mendel PDFДокумент5 страницVariações Das Leis de Mendel PDFmarceloОценок пока нет
- Exercicios de Respiração Celular e FermentaçãoДокумент3 страницыExercicios de Respiração Celular e FermentaçãoLara Coutinho100% (1)
- Hereditariedade: a transmissão de característicasДокумент14 страницHereditariedade: a transmissão de característicasMariana MargaridoОценок пока нет
- Leis de Mendel e conceitos fundamentais da herança genéticaДокумент72 страницыLeis de Mendel e conceitos fundamentais da herança genéticaDiego AlarconОценок пока нет
- A HereditariedadeДокумент10 страницA Hereditariedadejomar_marjoОценок пока нет
- Biologia 12o - Transmissão de Características GenéticasДокумент6 страницBiologia 12o - Transmissão de Características Genéticaspaula 25671Оценок пока нет
- Manual de Nutrição - Capítulo 4Документ6 страницManual de Nutrição - Capítulo 4Rafa DuarteОценок пока нет
- Manual de Nutrição - Capítulo 4Документ6 страницManual de Nutrição - Capítulo 4Rafa DuarteОценок пока нет
- O Japão Do TiagoДокумент3 страницыO Japão Do TiagoMariana MargaridoОценок пока нет
- Introdução TeóricaДокумент2 страницыIntrodução TeóricaMariana MargaridoОценок пока нет
- Introdução TeóricaДокумент2 страницыIntrodução TeóricaMariana MargaridoОценок пока нет
- Cap-25 Emerg ClinicasДокумент12 страницCap-25 Emerg Clinicasapi-3704607100% (1)
- Movimentos Transmembranares e Transporte CelularДокумент17 страницMovimentos Transmembranares e Transporte CelularMariana MargaridoОценок пока нет
- Regulação Genética, Mutações e CancroДокумент4 страницыRegulação Genética, Mutações e CancroMariana MargaridoОценок пока нет
- Camões Épico e LíricoДокумент3 страницыCamões Épico e LíricoMariana MargaridoОценок пока нет
- Cesario VerdeДокумент3 страницыCesario VerdeLucas SilvaОценок пока нет
- 12 ff4Документ1 страница12 ff4Mariana MargaridoОценок пока нет
- Relatório de QuímicaДокумент10 страницRelatório de QuímicaMariana MargaridoОценок пока нет
- 1 Parabéns Espero Que Já Saibas Que Te Adoro Muitoooooooo.. e Que Já Não Si Viver Sem Te Der Ao Meu Lado Sempre Que Tenho Algum StressДокумент1 страница1 Parabéns Espero Que Já Saibas Que Te Adoro Muitoooooooo.. e Que Já Não Si Viver Sem Te Der Ao Meu Lado Sempre Que Tenho Algum StressMariana MargaridoОценок пока нет
- RecursosДокумент28 страницRecursosMariana MargaridoОценок пока нет
- Teste LPДокумент10 страницTeste LPaldrabiceОценок пока нет
- Genética Humana AnáliseДокумент5 страницGenética Humana AnáliseMariana MargaridoОценок пока нет
- SANTILLANA - MAT12 FichaTrabalho - 08Документ2 страницыSANTILLANA - MAT12 FichaTrabalho - 08explimatОценок пока нет
- O Ano da Morte de Ricardo Reis de José SaramagoДокумент18 страницO Ano da Morte de Ricardo Reis de José SaramagoMariana MargaridoОценок пока нет
- FichaTrabalho MatematicaAДокумент3 страницыFichaTrabalho MatematicaANanda CostaОценок пока нет
- Hereditariedade: a transmissão de característicasДокумент14 страницHereditariedade: a transmissão de característicasMariana MargaridoОценок пока нет
- Estrutura Da Obra Obra Constituída Por 44 Poemas:: Mensagem - Análise de TextosДокумент5 страницEstrutura Da Obra Obra Constituída Por 44 Poemas:: Mensagem - Análise de TextosMariana MargaridoОценок пока нет
- O Ano da Morte de Ricardo Reis de José SaramagoДокумент18 страницO Ano da Morte de Ricardo Reis de José SaramagoMariana MargaridoОценок пока нет
- Analise de Poemas de "Mensagem" de Fernando PessoaДокумент62 страницыAnalise de Poemas de "Mensagem" de Fernando PessoaFlaviaRamalhaoОценок пока нет
- Analise de Poemas de "Mensagem" de Fernando PessoaДокумент62 страницыAnalise de Poemas de "Mensagem" de Fernando PessoaFlaviaRamalhaoОценок пока нет
- Dieta para Hipertrofia em 3 PassosДокумент23 страницыDieta para Hipertrofia em 3 PassosDinarte DuarteОценок пока нет
- Diretrizes EcocardiografiaДокумент43 страницыDiretrizes EcocardiografiaJuan NatãОценок пока нет
- Apostila de AnatomiaДокумент156 страницApostila de AnatomiaAnonymous CU1C9HwXОценок пока нет
- É Tido Como HemorragiaДокумент3 страницыÉ Tido Como HemorragiaGabrielОценок пока нет
- Intervenção com criança disléxicaДокумент114 страницIntervenção com criança disléxicaCláudia Oliveira100% (1)
- 04 NoçΣes de Anatomia e Fisiologia HumanaДокумент57 страниц04 NoçΣes de Anatomia e Fisiologia HumanaIsabella MartiniОценок пока нет
- Aula 1 - Farmacologia Do Sistema CardiovascularДокумент37 страницAula 1 - Farmacologia Do Sistema CardiovascularIsabelly EstefanyОценок пока нет
- Texto - Imunologia Da Mucosa OralДокумент20 страницTexto - Imunologia Da Mucosa OralLuana SouzaОценок пока нет
- Questionário de Rastreamento Metabólico3Документ3 страницыQuestionário de Rastreamento Metabólico3João Paulo Ramos Oliveira100% (3)
- Alimentação para Hiperuricémia e GotaДокумент2 страницыAlimentação para Hiperuricémia e GotaMarcia Rodrigues0% (1)
- Ação Do Ácido Salicílico Na Acne VulgarДокумент174 страницыAção Do Ácido Salicílico Na Acne VulgarAna Paula Pereira CunhaОценок пока нет
- Avaliação de Recuperação de Ciências - 7º AnoДокумент6 страницAvaliação de Recuperação de Ciências - 7º AnoBruno FerreiraОценок пока нет
- Protocolo odontológico para quimio e radioterapiaДокумент47 страницProtocolo odontológico para quimio e radioterapiaClaudio Luis VenturiniОценок пока нет
- Questões OdontopediatriaДокумент6 страницQuestões OdontopediatriaPatricia CorreaОценок пока нет
- Sistema CirculatórioДокумент7 страницSistema CirculatórioGheter CostaОценок пока нет
- Metilxantinas em plantasДокумент2 страницыMetilxantinas em plantasLuciana CapetiniОценок пока нет
- Ensino Secundário Regular - Biologia e Geologia 10o anoДокумент2 страницыEnsino Secundário Regular - Biologia e Geologia 10o anoJoaoОценок пока нет
- Fatores de RetençãoДокумент7 страницFatores de RetençãoFernanda FariaОценок пока нет
- Tabela de HonorariosДокумент5 страницTabela de HonorariosGeorgia Leandro SalernoОценок пока нет
- Olfação e Gustação 28 Abr 2023 Alex Kors Vidsiunas-1Документ18 страницOlfação e Gustação 28 Abr 2023 Alex Kors Vidsiunas-1minato.namikaze2901Оценок пока нет
- Revisão CitoplasmaДокумент3 страницыRevisão CitoplasmaClarice Agudo MenezesОценок пока нет
- Abreviaturas, Símbolos e Siglas Utilizadas PDFДокумент2 страницыAbreviaturas, Símbolos e Siglas Utilizadas PDFPedro SantosОценок пока нет
- Arte Fisiologia Vegetal PDFДокумент82 страницыArte Fisiologia Vegetal PDFClebersonRianОценок пока нет
- Aula Prática CinésioterapiaДокумент10 страницAula Prática CinésioterapiaGabi DinizОценок пока нет
- Arividade Contraste Julio - MoisesДокумент25 страницArividade Contraste Julio - Moisessfernanda.dutraОценок пока нет