Вам также может понравиться
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceОт EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceРейтинг: 4 из 5 звезд4/5 (895)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeОт EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeРейтинг: 4 из 5 звезд4/5 (5794)
- The Yellow House: A Memoir (2019 National Book Award Winner)От EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Рейтинг: 4 из 5 звезд4/5 (98)
- The Little Book of Hygge: Danish Secrets to Happy LivingОт EverandThe Little Book of Hygge: Danish Secrets to Happy LivingРейтинг: 3.5 из 5 звезд3.5/5 (400)
- Never Split the Difference: Negotiating As If Your Life Depended On ItОт EverandNever Split the Difference: Negotiating As If Your Life Depended On ItРейтинг: 4.5 из 5 звезд4.5/5 (838)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureОт EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureРейтинг: 4.5 из 5 звезд4.5/5 (474)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryОт EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryРейтинг: 3.5 из 5 звезд3.5/5 (231)
- The Emperor of All Maladies: A Biography of CancerОт EverandThe Emperor of All Maladies: A Biography of CancerРейтинг: 4.5 из 5 звезд4.5/5 (271)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaОт EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaРейтинг: 4.5 из 5 звезд4.5/5 (266)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersОт EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersРейтинг: 4.5 из 5 звезд4.5/5 (345)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyОт EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyРейтинг: 3.5 из 5 звезд3.5/5 (2259)
- Team of Rivals: The Political Genius of Abraham LincolnОт EverandTeam of Rivals: The Political Genius of Abraham LincolnРейтинг: 4.5 из 5 звезд4.5/5 (234)
- The Unwinding: An Inner History of the New AmericaОт EverandThe Unwinding: An Inner History of the New AmericaРейтинг: 4 из 5 звезд4/5 (45)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreОт EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreРейтинг: 4 из 5 звезд4/5 (1090)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)От EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Рейтинг: 4.5 из 5 звезд4.5/5 (121)
- Hemoglobin A1cДокумент3 страницыHemoglobin A1cFalah DakkaОценок пока нет
- L17 L18 Lipids 2013 14Документ23 страницыL17 L18 Lipids 2013 14Pejvak KHorram DinОценок пока нет
- MGISP-960: Compatible Kits For Different ConfigurationsДокумент2 страницыMGISP-960: Compatible Kits For Different ConfigurationshgfvvgffОценок пока нет
- Kinetics of Immobilized EnzymeДокумент10 страницKinetics of Immobilized EnzymeHari KumarОценок пока нет
- CarbohydratesДокумент51 страницаCarbohydratesYunita RachmawatiОценок пока нет
- Pcog LipidsДокумент3 страницыPcog LipidsCiara FajardoОценок пока нет
- Aspirin and Its Binding Sites: Week 3-4 Chris CarsonДокумент12 страницAspirin and Its Binding Sites: Week 3-4 Chris CarsonKit CarsonОценок пока нет
- Structure of LipidДокумент11 страницStructure of LipidAfrah Alatas100% (1)
- Sankar Et Al., 2013Документ6 страницSankar Et Al., 2013Natasha MaharaniОценок пока нет
- Cell Physiology MCQДокумент20 страницCell Physiology MCQRD DaskaОценок пока нет
- Khan Academy Carbohydrates QuestionsДокумент3 страницыKhan Academy Carbohydrates QuestionsLoraОценок пока нет
- Vit & Enzyme (Group3) Final OutlineДокумент4 страницыVit & Enzyme (Group3) Final OutlineJohn Mark LibreОценок пока нет
- BioChem LecДокумент21 страницаBioChem LecAbby Dimalaluan OquendoОценок пока нет
- Lec - Chemical MessengersДокумент5 страницLec - Chemical MessengersSim QuiambaoОценок пока нет
- JELS Vol 2 No 2 2012 PDFДокумент57 страницJELS Vol 2 No 2 2012 PDFErika ViyanaОценок пока нет
- Deoxyribonucleic AcidДокумент36 страницDeoxyribonucleic AcidAlex LlanoОценок пока нет
- 2 Introduction - To - Detergent - Enzymes PDFДокумент45 страниц2 Introduction - To - Detergent - Enzymes PDFtmlОценок пока нет
- Multiple Choice Questions: BiotechnologyДокумент98 страницMultiple Choice Questions: BiotechnologyJing Li100% (1)
- Copper CVD Final2Документ8 страницCopper CVD Final2api-241644265Оценок пока нет
- Final Estimation of SGOTДокумент3 страницыFinal Estimation of SGOTshibsankar rakshitОценок пока нет
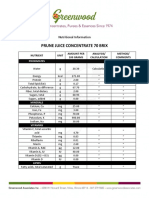
- Prune Juice Concentrate 70 Brix Nutritional InformationДокумент2 страницыPrune Juice Concentrate 70 Brix Nutritional Informationborn2dive 9702Оценок пока нет
- Illumina Dye SequencingДокумент5 страницIllumina Dye SequencingNguyen TaОценок пока нет
- ICD 10 Diagnosis GiziДокумент2 страницыICD 10 Diagnosis Gizigizi cgcОценок пока нет
- Fundamentals of EnzymologyДокумент3 страницыFundamentals of EnzymologySuryaditya TanejaОценок пока нет
- AnticancerДокумент78 страницAnticancerRajkishor GogoiОценок пока нет
- Rna PDFДокумент12 страницRna PDFBelegurthОценок пока нет
- Antioxidants in Biology and Medicine - Essentials, Advances, and Clinical ApplicationsДокумент431 страницаAntioxidants in Biology and Medicine - Essentials, Advances, and Clinical Applicationsnrh2005100% (1)
- Prolonging Healthy Aging - Longevity Vitamins and Proteins - PNASДокумент16 страницProlonging Healthy Aging - Longevity Vitamins and Proteins - PNASgodolfintorentОценок пока нет
- General Biology 1 Module FINALSДокумент12 страницGeneral Biology 1 Module FINALSKenneth Vince AlonzoОценок пока нет
- BiologyДокумент24 страницыBiologyTom KiloitОценок пока нет