Вам также может понравиться
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeОт EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeРейтинг: 4 из 5 звезд4/5 (5794)
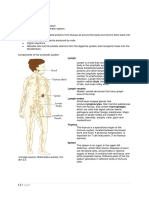
- Lymphatic SystemДокумент10 страницLymphatic SystemPasipanodya MuzendaОценок пока нет
- Nut2601 2016 10 e 1Документ5 страницNut2601 2016 10 e 1Pasipanodya MuzendaОценок пока нет
- Bmi2603 2016 6 e 1Документ2 страницыBmi2603 2016 6 e 1Pasipanodya MuzendaОценок пока нет
- Tutorial Letter 101/3/2019: Molecular Cell BiologyДокумент11 страницTutorial Letter 101/3/2019: Molecular Cell BiologyPasipanodya MuzendaОценок пока нет
- Study Unit No Examination Date Time DurationДокумент1 страницаStudy Unit No Examination Date Time DurationPasipanodya MuzendaОценок пока нет
- What Are Stem CellsДокумент8 страницWhat Are Stem CellsPasipanodya MuzendaОценок пока нет
- Memo For OctoberДокумент5 страницMemo For OctoberPasipanodya MuzendaОценок пока нет
- Sta1510 2016 6 e 1Документ23 страницыSta1510 2016 6 e 1Pasipanodya Muzenda100% (2)
- Bachelor of Science Biomedical ScienceДокумент1 страницаBachelor of Science Biomedical SciencePasipanodya Muzenda100% (1)
- Blg1502 Exam Prep Doc - Part 1&2Документ18 страницBlg1502 Exam Prep Doc - Part 1&2Pasipanodya MuzendaОценок пока нет
- BLG1502 2017 6 e 1Документ5 страницBLG1502 2017 6 e 1Pasipanodya MuzendaОценок пока нет
- My Notes For Blg1502!1!1Документ10 страницMy Notes For Blg1502!1!1Pasipanodya MuzendaОценок пока нет
- STA1510 STA1610 2017 TL Part I Questions and SolutionsДокумент33 страницыSTA1510 STA1610 2017 TL Part I Questions and SolutionsPasipanodya Muzenda60% (5)
- Solutions To Assignment 1Документ7 страницSolutions To Assignment 1Pasipanodya MuzendaОценок пока нет
- HCL Pyridine Cro (PCC) CH CL Oh O H: Key Oxidizing Agents KeyДокумент1 страницаHCL Pyridine Cro (PCC) CH CL Oh O H: Key Oxidizing Agents KeyPasipanodya MuzendaОценок пока нет
- E2 MechanismДокумент2 страницыE2 MechanismPasipanodya MuzendaОценок пока нет
- Oxidizing Agents Draw The Organic Product From Each ReactionДокумент1 страницаOxidizing Agents Draw The Organic Product From Each ReactionPasipanodya MuzendaОценок пока нет
- BLG1502 Assignment No 2 S2 2015 MemoДокумент7 страницBLG1502 Assignment No 2 S2 2015 MemoPasipanodya Muzenda80% (5)
- Che1501 2015 10 e 1Документ18 страницChe1501 2015 10 e 1Pasipanodya MuzendaОценок пока нет
- Factors Affecting Body CompositionДокумент2 страницыFactors Affecting Body CompositionPasipanodya MuzendaОценок пока нет
- Che1501 2016 6 e 1Документ17 страницChe1501 2016 6 e 1Pasipanodya MuzendaОценок пока нет
- 101 2017 3 B PDFДокумент18 страниц101 2017 3 B PDFPasipanodya MuzendaОценок пока нет
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceОт EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceРейтинг: 4 из 5 звезд4/5 (895)
- The Yellow House: A Memoir (2019 National Book Award Winner)От EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Рейтинг: 4 из 5 звезд4/5 (98)
- The Little Book of Hygge: Danish Secrets to Happy LivingОт EverandThe Little Book of Hygge: Danish Secrets to Happy LivingРейтинг: 3.5 из 5 звезд3.5/5 (400)
- The Emperor of All Maladies: A Biography of CancerОт EverandThe Emperor of All Maladies: A Biography of CancerРейтинг: 4.5 из 5 звезд4.5/5 (271)
- Never Split the Difference: Negotiating As If Your Life Depended On ItОт EverandNever Split the Difference: Negotiating As If Your Life Depended On ItРейтинг: 4.5 из 5 звезд4.5/5 (838)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyОт EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyРейтинг: 3.5 из 5 звезд3.5/5 (2259)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureОт EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureРейтинг: 4.5 из 5 звезд4.5/5 (474)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryОт EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryРейтинг: 3.5 из 5 звезд3.5/5 (231)
- Team of Rivals: The Political Genius of Abraham LincolnОт EverandTeam of Rivals: The Political Genius of Abraham LincolnРейтинг: 4.5 из 5 звезд4.5/5 (234)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaОт EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaРейтинг: 4.5 из 5 звезд4.5/5 (266)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersОт EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersРейтинг: 4.5 из 5 звезд4.5/5 (345)
- The Unwinding: An Inner History of the New AmericaОт EverandThe Unwinding: An Inner History of the New AmericaРейтинг: 4 из 5 звезд4/5 (45)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreОт EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreРейтинг: 4 из 5 звезд4/5 (1090)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)От EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Рейтинг: 4.5 из 5 звезд4.5/5 (121)
- Problem ManagementДокумент33 страницыProblem Managementdhirajsatyam98982285Оценок пока нет
- 01ESS - Introducing Siebel ApplicationsДокумент24 страницы01ESS - Introducing Siebel ApplicationsRajaОценок пока нет
- GearsДокумент14 страницGearsZulhilmi Chik TakОценок пока нет
- Poet Forugh Farrokhzad in World Poetry PДокумент3 страницыPoet Forugh Farrokhzad in World Poetry Pkarla telloОценок пока нет
- Mathsnacks05 InfiniteДокумент1 страницаMathsnacks05 Infiniteburkard.polsterОценок пока нет
- Work Teams and GroupsДокумент6 страницWork Teams and GroupsFides AvendanОценок пока нет
- Affirmative (Afirmativa) Long Form Short Form PortuguêsДокумент3 страницыAffirmative (Afirmativa) Long Form Short Form PortuguêsAnitaYangОценок пока нет
- Approach To Vaginal Discharge in ChildrenДокумент12 страницApproach To Vaginal Discharge in ChildrensujataОценок пока нет
- A Passage To AfricaДокумент25 страницA Passage To AfricaJames Reinz100% (2)
- Counselling and PsychotherapyДокумент12 страницCounselling and PsychotherapyD.Sreenivasa ReddyОценок пока нет
- Percy Bysshe ShelleyДокумент20 страницPercy Bysshe Shelleynishat_haider_2100% (1)
- Best-First SearchДокумент2 страницыBest-First Searchgabby209Оценок пока нет
- GRADE 1 To 12 Daily Lesson LOG: TLE6AG-Oc-3-1.3.3Документ7 страницGRADE 1 To 12 Daily Lesson LOG: TLE6AG-Oc-3-1.3.3Roxanne Pia FlorentinoОценок пока нет
- Validator in JSFДокумент5 страницValidator in JSFvinh_kakaОценок пока нет
- Serological and Molecular DiagnosisДокумент9 страницSerological and Molecular DiagnosisPAIRAT, Ella Joy M.Оценок пока нет
- CEI and C4C Integration in 1602: Software Design DescriptionДокумент44 страницыCEI and C4C Integration in 1602: Software Design Descriptionpkumar2288Оценок пока нет
- Peri Operative Nursing ManagementДокумент19 страницPeri Operative Nursing ManagementSabina KontehОценок пока нет
- Glickman - The Jewish White Slavery Trade (2000)Документ152 страницыGlickman - The Jewish White Slavery Trade (2000)Alrik G. HamerОценок пока нет
- 2023-Tutorial 02Документ6 страниц2023-Tutorial 02chyhyhyОценок пока нет
- Department of Chemistry Ramakrishna Mission V. C. College, RaharaДокумент16 страницDepartment of Chemistry Ramakrishna Mission V. C. College, RaharaSubhro ChatterjeeОценок пока нет
- A2B1 Unit 11bДокумент2 страницыA2B1 Unit 11bTheToan.Оценок пока нет
- Feline Neonatal IsoerythrolysisДокумент18 страницFeline Neonatal IsoerythrolysisPaunas JoshiОценок пока нет
- Final Presentation BANK OF BARODA 1Документ8 страницFinal Presentation BANK OF BARODA 1Pooja GoyalОценок пока нет
- Producto Académico #2: Inglés Profesional 2Документ2 страницыProducto Académico #2: Inglés Profesional 2fredy carpioОценок пока нет
- Marketing PlanДокумент41 страницаMarketing PlanMark AbainzaОценок пока нет
- Enunciado de La Pregunta: Finalizado Se Puntúa 1.00 Sobre 1.00Документ9 страницEnunciado de La Pregunta: Finalizado Se Puntúa 1.00 Sobre 1.00Samuel MojicaОценок пока нет
- Leg Res Cases 4Документ97 страницLeg Res Cases 4acheron_pОценок пока нет
- ObliCon Digests PDFДокумент48 страницObliCon Digests PDFvictoria pepitoОценок пока нет
- Annexure 8: Medical Certificate (To Be Issued by A Registered Medical Practitioner) General ExpectationsДокумент1 страницаAnnexure 8: Medical Certificate (To Be Issued by A Registered Medical Practitioner) General ExpectationsMannepalli RamakrishnaОценок пока нет
- Re CrystallizationДокумент25 страницRe CrystallizationMarol CerdaОценок пока нет