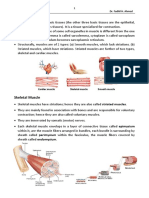
Вам также может понравиться
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryОт EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryРейтинг: 3.5 из 5 звезд3.5/5 (231)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)От EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Рейтинг: 4.5 из 5 звезд4.5/5 (119)
- Never Split the Difference: Negotiating As If Your Life Depended On ItОт EverandNever Split the Difference: Negotiating As If Your Life Depended On ItРейтинг: 4.5 из 5 звезд4.5/5 (838)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaОт EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaРейтинг: 4.5 из 5 звезд4.5/5 (265)
- The Little Book of Hygge: Danish Secrets to Happy LivingОт EverandThe Little Book of Hygge: Danish Secrets to Happy LivingРейтинг: 3.5 из 5 звезд3.5/5 (399)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyОт EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyРейтинг: 3.5 из 5 звезд3.5/5 (2219)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeОт EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeРейтинг: 4 из 5 звезд4/5 (5794)
- Team of Rivals: The Political Genius of Abraham LincolnОт EverandTeam of Rivals: The Political Genius of Abraham LincolnРейтинг: 4.5 из 5 звезд4.5/5 (234)
- The Emperor of All Maladies: A Biography of CancerОт EverandThe Emperor of All Maladies: A Biography of CancerРейтинг: 4.5 из 5 звезд4.5/5 (271)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreОт EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreРейтинг: 4 из 5 звезд4/5 (1090)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersОт EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersРейтинг: 4.5 из 5 звезд4.5/5 (344)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceОт EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceРейтинг: 4 из 5 звезд4/5 (890)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureОт EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureРейтинг: 4.5 из 5 звезд4.5/5 (474)
- The Unwinding: An Inner History of the New AmericaОт EverandThe Unwinding: An Inner History of the New AmericaРейтинг: 4 из 5 звезд4/5 (45)
- The Yellow House: A Memoir (2019 National Book Award Winner)От EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Рейтинг: 4 из 5 звезд4/5 (98)
- Excitable Tissue PhysiologyДокумент13 страницExcitable Tissue PhysiologyAlbert Che33% (3)
- Ethem Murat Arsava (Eds.) - Nutrition in Neurologic Disorders - A Practical Guide (2017, Springer International Publishing)Документ209 страницEthem Murat Arsava (Eds.) - Nutrition in Neurologic Disorders - A Practical Guide (2017, Springer International Publishing)riddhiОценок пока нет
- POGIL KeyДокумент2 страницыPOGIL KeyfiueОценок пока нет
- A Multiparametric Mri Score For Prostate Cancer Detection Performance in Patients With and Without Endorectal CoilДокумент8 страницA Multiparametric Mri Score For Prostate Cancer Detection Performance in Patients With and Without Endorectal CoilShira michaelОценок пока нет
- Highintensity Focused Ultrasound of Uterine Fibroids and Adenomyosis Maneuver Technique For BowelДокумент3 страницыHighintensity Focused Ultrasound of Uterine Fibroids and Adenomyosis Maneuver Technique For BowelShira michaelОценок пока нет
- Successful Retrieval of A Dislodged Zotarolimuseluting Coronary Stent in The Ascending AortaДокумент3 страницыSuccessful Retrieval of A Dislodged Zotarolimuseluting Coronary Stent in The Ascending AortaShira michaelОценок пока нет
- Interference With Creatinine Estimation by Impenem Cilastatin in Jaffes Kinetic MethodДокумент2 страницыInterference With Creatinine Estimation by Impenem Cilastatin in Jaffes Kinetic MethodShira michaelОценок пока нет
- Respiratory Function Following Onpump Versus Offpump Coronary Bypass Grafting SurgeryДокумент7 страницRespiratory Function Following Onpump Versus Offpump Coronary Bypass Grafting SurgeryShira michaelОценок пока нет
- Comparison of Endotracheal Cuff Pressure Measurements Before and After Nursing Care in Emergency Patients Pilot BalloonДокумент5 страницComparison of Endotracheal Cuff Pressure Measurements Before and After Nursing Care in Emergency Patients Pilot BalloonShira michaelОценок пока нет
- Effects of Exenatide On Both Fat Mass Loss and Cardiometabolic Risk in Obese Subjects With Type 2 Diabetes MellitusДокумент7 страницEffects of Exenatide On Both Fat Mass Loss and Cardiometabolic Risk in Obese Subjects With Type 2 Diabetes MellitusShira michaelОценок пока нет
- A Clinical Study of Adjuvant Chemotherapy in Younger and Elder Rectal Cancer PatientsaДокумент12 страницA Clinical Study of Adjuvant Chemotherapy in Younger and Elder Rectal Cancer PatientsaShira michaelОценок пока нет
- Rheumatology To Hepatology Cross Talk An Evidence Based Update On The Treat To Target Strategy in Hepatitisc Related ExtДокумент9 страницRheumatology To Hepatology Cross Talk An Evidence Based Update On The Treat To Target Strategy in Hepatitisc Related ExtShira michaelОценок пока нет
- Onset Patterns of Chronic Fatigue Syndrome and Myalgic EncephalomyelitisДокумент30 страницOnset Patterns of Chronic Fatigue Syndrome and Myalgic EncephalomyelitisShira michaelОценок пока нет
- Comparison of Rigid and Deformable Image Registration Accuracy of The Liver During Longterm Transition After Proton BeamДокумент6 страницComparison of Rigid and Deformable Image Registration Accuracy of The Liver During Longterm Transition After Proton BeamShira michaelОценок пока нет
- The Role of History of Disease Factors in The Congenital Heart Disease in AdultsДокумент4 страницыThe Role of History of Disease Factors in The Congenital Heart Disease in AdultsShira michaelОценок пока нет
- Highintensity Focused Ultrasound of Uterine Fibroids and Adenomyosis Maneuver Technique For BowelДокумент3 страницыHighintensity Focused Ultrasound of Uterine Fibroids and Adenomyosis Maneuver Technique For BowelShira michaelОценок пока нет
- Quantitative Analysis On Literature of Working Memory On The Athlete in ChinaДокумент5 страницQuantitative Analysis On Literature of Working Memory On The Athlete in ChinaShira michaelОценок пока нет
- Should Plain Radiographs Persist or Be Replaced by Alternative Scans For Imaging of Acute Painful Nontraumatic AbdominalДокумент6 страницShould Plain Radiographs Persist or Be Replaced by Alternative Scans For Imaging of Acute Painful Nontraumatic AbdominalShira michaelОценок пока нет
- Successful Retrieval of A Dislodged Zotarolimuseluting Coronary Stent in The Ascending AortaДокумент3 страницыSuccessful Retrieval of A Dislodged Zotarolimuseluting Coronary Stent in The Ascending AortaShira michaelОценок пока нет
- Yatri R. Shah, Dhrubo Jyoti Sen and C N Patel - Schiff's Bases of Piperidone Derivative As Microbial Growth InhibitorsДокумент9 страницYatri R. Shah, Dhrubo Jyoti Sen and C N Patel - Schiff's Bases of Piperidone Derivative As Microbial Growth InhibitorsSmokeysamОценок пока нет
- Phytochemical and Antimicrobial Studies of Oroxylum Indicum ExtractsДокумент8 страницPhytochemical and Antimicrobial Studies of Oroxylum Indicum ExtractsShira michaelОценок пока нет
- Comparison of Endotracheal Cuff Pressure Measurements Before and After Nursing Care in Emergency Patients Pilot BalloonДокумент5 страницComparison of Endotracheal Cuff Pressure Measurements Before and After Nursing Care in Emergency Patients Pilot BalloonShira michaelОценок пока нет
- A Rare Complication of Acute Cholecystitis Leading To Perihepatic Abscess Gall Bladder PerforationДокумент3 страницыA Rare Complication of Acute Cholecystitis Leading To Perihepatic Abscess Gall Bladder PerforationShira michaelОценок пока нет
- 3 - Cells and TissuesДокумент20 страниц3 - Cells and TissuesTwelve Forty-fourОценок пока нет
- Agr122 Lab ReportДокумент12 страницAgr122 Lab ReportNur AthirahОценок пока нет
- Grade 12 NSC Life Sciences P2 (English) Preparatory Examination Possible AnswersДокумент14 страницGrade 12 NSC Life Sciences P2 (English) Preparatory Examination Possible Answersnthakotebza629Оценок пока нет
- Mascular TissueДокумент4 страницыMascular TissueFadhil Hussam AhmedОценок пока нет
- Cell anatomy and divisionДокумент4 страницыCell anatomy and divisionMarielle SaymanОценок пока нет
- Polysaccaride Krestin (PSK)Документ5 страницPolysaccaride Krestin (PSK)Andromeda AbrahamОценок пока нет
- Physioex Lab Report: Pre-Lab Quiz ResultsДокумент4 страницыPhysioex Lab Report: Pre-Lab Quiz ResultsNabila Azzahra0% (1)
- Cell Structure and TaxonomyДокумент50 страницCell Structure and TaxonomyCarl Elexer Cuyugan AnoОценок пока нет
- An Anti Apoptotic HEK293 Cell Line Provides A Robust and High Titer Platform For Transient Protein Expression in BioreactorsДокумент11 страницAn Anti Apoptotic HEK293 Cell Line Provides A Robust and High Titer Platform For Transient Protein Expression in BioreactorsAtrocitus RedОценок пока нет
- IGCSE Enzyme GuideДокумент28 страницIGCSE Enzyme GuideawanbioscienceОценок пока нет
- CDU BIOCHEMISTRY Proteomics Amino Acids WORKSHEETДокумент4 страницыCDU BIOCHEMISTRY Proteomics Amino Acids WORKSHEETKrisha Mae VillanuevaОценок пока нет
- The Role of Zinc in Antiviral Immunity: ReviewДокумент15 страницThe Role of Zinc in Antiviral Immunity: ReviewNandkumar ChinaiОценок пока нет
- 2012 Smarcal1 Human Molecular GeneticsДокумент867 страниц2012 Smarcal1 Human Molecular Geneticsrameshramyb100% (1)
- Metabolism of Carbohydrates, Lipids, Amino Acids and its Regulation Test QuestionsДокумент209 страницMetabolism of Carbohydrates, Lipids, Amino Acids and its Regulation Test Questionsninas1112Оценок пока нет
- Chakraborti-Das2020 Article DysbiosisOfOralMicrobiotaAndItДокумент12 страницChakraborti-Das2020 Article DysbiosisOfOralMicrobiotaAndItraunak81Оценок пока нет
- Cellular Membranes: Structures and FunctionsДокумент7 страницCellular Membranes: Structures and FunctionsSwasti Agnihotri100% (1)
- Oncology Pathology Tanuvas NotesДокумент17 страницOncology Pathology Tanuvas NotesNagesh NОценок пока нет
- The Reactome BookДокумент3 252 страницыThe Reactome BookA RoyОценок пока нет
- Embryonic Stem CellsДокумент3 страницыEmbryonic Stem CellsRajОценок пока нет
- Biology Notes: EnzymesДокумент7 страницBiology Notes: Enzymeslaiba amirОценок пока нет
- Cytokine Storm Response To COVID-19 VaccinationsДокумент3 страницыCytokine Storm Response To COVID-19 VaccinationsewewewОценок пока нет
- A. Competitive Inhibition: Table 3Документ1 страницаA. Competitive Inhibition: Table 3Tran TuanОценок пока нет
- Cell Lysis ProtocolДокумент5 страницCell Lysis ProtocolmnstnОценок пока нет
- Chapter 7. Elements of Host Protective Responses - ClinicalKeyДокумент12 страницChapter 7. Elements of Host Protective Responses - ClinicalKeyد. محمد سند راشد الحصينيОценок пока нет
- AP Bio Chapter 13 NotesДокумент5 страницAP Bio Chapter 13 NotesAndrew SongОценок пока нет
- RP - MicrosДокумент17 страницRP - MicrosNeel MutagiОценок пока нет
- Sinh Lý MáuДокумент4 страницыSinh Lý MáuJonh MikeОценок пока нет