Вам также может понравиться
- T3M3-Metabolismo de Los GlucidosДокумент10 страницT3M3-Metabolismo de Los GlucidosEdersón RojasОценок пока нет
- Metabolisimo Del Glucógeno Glucogenosis y GlucogenogenesisДокумент22 страницыMetabolisimo Del Glucógeno Glucogenosis y GlucogenogenesisAngela BaezОценок пока нет
- GluconeogénesisДокумент23 страницыGluconeogénesisMario Joseph Escajadillo PorrasОценок пока нет
- Monografia (Ta1) P.BДокумент6 страницMonografia (Ta1) P.BDaniel RivasОценок пока нет
- Informe de Bioquimica Semana - 5Документ9 страницInforme de Bioquimica Semana - 5Ray CarSoОценок пока нет
- GLUCOSAДокумент10 страницGLUCOSAIvan DiazОценок пока нет
- GLUCONEOGÉNESISДокумент13 страницGLUCONEOGÉNESISLismar Katherin RamirezОценок пока нет
- GLUCOGENOДокумент18 страницGLUCOGENOUn pedacito de CieloОценок пока нет
- GLUCOGENÓLISISДокумент45 страницGLUCOGENÓLISISCarlos PlascenciaОценок пока нет
- Glucolisis Fmed PDFДокумент24 страницыGlucolisis Fmed PDFRamón BrunoОценок пока нет
- Informe de Lab N°8 - IndividualДокумент8 страницInforme de Lab N°8 - IndividualMiluzka Achata CulqueОценок пока нет
- Nombre: Registro: Grupo: Materia:: Marisol Nava Cervantes 217173837 B1 Metabolismo LДокумент4 страницыNombre: Registro: Grupo: Materia:: Marisol Nava Cervantes 217173837 B1 Metabolismo LNava Cervantes MarisolОценок пока нет
- P1 A.B.Документ7 страницP1 A.B.KIRSA BARBARA TACO ROQUEОценок пока нет
- Func. Hepat. Zarkin Reynoso Maria FernandaДокумент1 страницаFunc. Hepat. Zarkin Reynoso Maria FernandaMarifer ZarkinОценок пока нет
- Teórico 10 Dra Cymeryng - Texto Regulación Del Metabolismo de Glúcidos - 2018 RESALTADOДокумент21 страницаTeórico 10 Dra Cymeryng - Texto Regulación Del Metabolismo de Glúcidos - 2018 RESALTADOvivi soriaОценок пока нет
- Ludica Gluconeogenesis 1.Документ34 страницыLudica Gluconeogenesis 1.Pricila Pupo PuertasОценок пока нет
- Resumen de La Integración MetabólicaДокумент9 страницResumen de La Integración MetabólicaJuan Carlos Caredano PalaciosОценок пока нет
- Regulación Del Metabolismo de La Glucosa Desde Una Perspectiva Hepática.Документ6 страницRegulación Del Metabolismo de La Glucosa Desde Una Perspectiva Hepática.andreaОценок пока нет
- Bioquímica de Los Carbohidrato PDFДокумент6 страницBioquímica de Los Carbohidrato PDFHeinz Leif Guibovich EspinoОценок пока нет
- Aislamiento Cuantificacion Del Glucogeno Hepatico y MuscularДокумент8 страницAislamiento Cuantificacion Del Glucogeno Hepatico y MuscularIsaias MacuriiОценок пока нет
- Practica de GlucogenoДокумент11 страницPractica de GlucogenoKarim Milushka Ayala75% (4)
- Práctica1 Cuestionario FernandezДокумент6 страницPráctica1 Cuestionario Fernandezyadira fernandezОценок пока нет
- La GlucosaДокумент24 страницыLa GlucosaAngela Santiago OrtegaОценок пока нет
- Practica #6Документ32 страницыPractica #6Eder MariñasОценок пока нет
- Bioquimica y Su Lab - 6Документ5 страницBioquimica y Su Lab - 6Juan Junior Ullola RamirezОценок пока нет
- Glucogénesis y GlucogenólisisДокумент10 страницGlucogénesis y GlucogenólisisKathia Joselin Constantino MasОценок пока нет
- Bme U1 Ea KacmДокумент7 страницBme U1 Ea KacmKacoma MartínezОценок пока нет
- Informe 4 Bioquímica Seminario - Grupo 07Документ12 страницInforme 4 Bioquímica Seminario - Grupo 07Emery Sofía C. D. Bustamante0% (1)
- Bioquimica-Elizabeth y KarenДокумент16 страницBioquimica-Elizabeth y KarenDayana Elizabeth Gordillo NavarreteОценок пока нет
- Metabolismo de Los CarbohidratosДокумент51 страницаMetabolismo de Los CarbohidratosMaria Angelica Santillan Carrasco0% (1)
- Seminario Fructosa BioquimicaДокумент26 страницSeminario Fructosa BioquimicaMARISOL OYARCE LLAJAОценок пока нет
- Tarea 4 - BM - KHP PDFДокумент11 страницTarea 4 - BM - KHP PDFKenia HernandezОценок пока нет
- Cuestionario Metabolismo Del GlucogenoДокумент3 страницыCuestionario Metabolismo Del GlucogenoMaria Jose Mercado BarretoОценок пока нет
- Teórico 10 - Regulación Del Metabolismo de Glúcidos - 2021 - RectificadoДокумент24 страницыTeórico 10 - Regulación Del Metabolismo de Glúcidos - 2021 - RectificadoVirginia GalloОценок пока нет
- Seminario 7 - Metabolismo de Glúcidos I. Glucólisis, Gluconeogénesis y Vía de Las Pentosas - 2015Документ35 страницSeminario 7 - Metabolismo de Glúcidos I. Glucólisis, Gluconeogénesis y Vía de Las Pentosas - 2015Sujei ChavezОценок пока нет
- Clases Teoricas Metabolismo de Los CarbohidratosДокумент42 страницыClases Teoricas Metabolismo de Los CarbohidratosJose Luis BustamanteОценок пока нет
- Gluconeogénesis y Control de La Glucosa SanguíneaДокумент3 страницыGluconeogénesis y Control de La Glucosa SanguíneaDaniela Rosero100% (1)
- GlucogenólisisДокумент7 страницGlucogenólisisサンティアゴ クリスティアンОценок пока нет
- Glucógeno Muscular Vs Glucógeno HepáticoДокумент10 страницGlucógeno Muscular Vs Glucógeno HepáticoAdriana GonzalezОценок пока нет
- Practica GlucogenoДокумент25 страницPractica GlucogenoMiguel Angel Rodas Herrera75% (24)
- Principales Formas de Regulacion Del MetabolismoДокумент13 страницPrincipales Formas de Regulacion Del MetabolismoWendy BarrazaОценок пока нет
- Glucogénesis Bioquimica ClinicaДокумент12 страницGlucogénesis Bioquimica ClinicaDavid AbregoОценок пока нет
- INFORME LABORATORIO GLUCOGENO - Manuela Correa RíosДокумент8 страницINFORME LABORATORIO GLUCOGENO - Manuela Correa RíosManuela Correa RiosОценок пока нет
- Monografia BiologiaДокумент9 страницMonografia BiologiaJoseph Mamani IngaluqueОценок пока нет
- Taller #4Документ6 страницTaller #4Andy MoreiraОценок пока нет
- Guias 9, 10, 11 Bioquimica LUIS DONADOДокумент7 страницGuias 9, 10, 11 Bioquimica LUIS DONADOLuis Carlos Donado del AguilaОценок пока нет
- Sandoval - José - Investigación 1Документ4 страницыSandoval - José - Investigación 1MiguelОценок пока нет
- Glicemia Cuestionario GuevaraДокумент2 страницыGlicemia Cuestionario Guevarajhossgm8Оценок пока нет
- Metabolismo Carbohidratos (Teoria)Документ20 страницMetabolismo Carbohidratos (Teoria)henryОценок пока нет
- Lab 3 GlicemiaДокумент7 страницLab 3 GlicemiaDiana MendezОценок пока нет
- Metabolismo de CHOДокумент15 страницMetabolismo de CHOoscar giraldoОценок пока нет
- Glucolisis y Glucogenesis ALEJANDROДокумент5 страницGlucolisis y Glucogenesis ALEJANDROYotdan martinezОценок пока нет
- TALLER N 3 BioquimicaДокумент6 страницTALLER N 3 Bioquimicaadelina hernandez100% (1)
- Relaciones Metabolicas Tisulares enДокумент30 страницRelaciones Metabolicas Tisulares enSimón WitonОценок пока нет
- Metabolismo CHO, Lípidos y ProteínasДокумент23 страницыMetabolismo CHO, Lípidos y ProteínasAdrian MorganОценок пока нет
- INFORME S4 ESO26 Bioquímica Seminario VVVДокумент11 страницINFORME S4 ESO26 Bioquímica Seminario VVVCoraine Chávez CórdovaОценок пока нет
- Unidad 3. Metabolismo de CarbohidratosДокумент15 страницUnidad 3. Metabolismo de CarbohidratosCristiana Auxiliadora Fonseca MayorgaОценок пока нет
- 4-Metabolismo Carbohidratos P-1Документ58 страниц4-Metabolismo Carbohidratos P-1Nar JovОценок пока нет
- TALLER - No. - 1 - de - BIOQUIMICA - 2021 - 2 - DEFДокумент28 страницTALLER - No. - 1 - de - BIOQUIMICA - 2021 - 2 - DEFlaura pardoОценок пока нет
- Caso Clínico de Streptococcus PyogenesДокумент3 страницыCaso Clínico de Streptococcus PyogenesMario Bacilio Escobedo100% (1)
- Diabetes Tipo 2 en Niños y AdolescentesДокумент21 страницаDiabetes Tipo 2 en Niños y AdolescentesMario Bacilio EscobedoОценок пока нет
- CASO CLINICO Haemophilus InfluenzaeДокумент3 страницыCASO CLINICO Haemophilus InfluenzaeMario Bacilio Escobedo100% (1)
- CASO CLINICO DE Corynebacterium DiphtheriaeДокумент6 страницCASO CLINICO DE Corynebacterium DiphtheriaeMario Bacilio EscobedoОценок пока нет
- Efectos Del Glucagón y La Insulina en La Concentración Plasmática de Glucosa en Los Deportistas.Документ14 страницEfectos Del Glucagón y La Insulina en La Concentración Plasmática de Glucosa en Los Deportistas.Mario Bacilio EscobedoОценок пока нет
- Reglamento Del LaboratorioДокумент2 страницыReglamento Del LaboratorioMario Bacilio EscobedoОценок пока нет
- EsquizofreniaДокумент77 страницEsquizofreniaMario Bacilio EscobedoОценок пока нет
- Agentes AntisépticossДокумент1 страницаAgentes AntisépticossMario Bacilio EscobedoОценок пока нет
- NORMA Oficial Mexicana NOM-028-SSA2-1999, para La Prevención, Tratamiento y Control de Las Adicciones.Документ27 страницNORMA Oficial Mexicana NOM-028-SSA2-1999, para La Prevención, Tratamiento y Control de Las Adicciones.Mario Bacilio EscobedoОценок пока нет
- Agentes AntisépticossДокумент1 страницаAgentes AntisépticossMario Bacilio EscobedoОценок пока нет
- Agentes AntisépticossДокумент1 страницаAgentes AntisépticossMario Bacilio EscobedoОценок пока нет
- Precauciones UniversalesДокумент18 страницPrecauciones UniversalesMario Bacilio Escobedo0% (1)
- Resumen de SociomedicaДокумент3 страницыResumen de SociomedicaMario Bacilio EscobedoОценок пока нет
- Embriologia Humana Mariana RojasДокумент54 страницыEmbriologia Humana Mariana RojasFrancisca Valentina Cárcamo UlloaОценок пока нет
- ASpectos Psicologicos de La Hospitalizacion InfantilДокумент6 страницASpectos Psicologicos de La Hospitalizacion InfantilMario Bacilio EscobedoОценок пока нет
- Manual de DeportesДокумент7 страницManual de DeportesMario Bacilio EscobedoОценок пока нет
- Cómo Iniciar Una Conversación Con Cualquier Persona y en Cualquier MomentoДокумент12 страницCómo Iniciar Una Conversación Con Cualquier Persona y en Cualquier MomentoFRANCISCO JAVIER SALAZAR NUÑEZ100% (3)
- Como Hablar Con Una MujerДокумент9 страницComo Hablar Con Una MujerMario Bacilio Escobedo100% (2)
- Ensayo de Antropologia MedicaДокумент5 страницEnsayo de Antropologia MedicaMario Bacilio EscobedoОценок пока нет
- ¿Nosotros Utilizamos Sólo El 10% de Nuestro Cerebro?Документ1 страница¿Nosotros Utilizamos Sólo El 10% de Nuestro Cerebro?Mario Bacilio EscobedoОценок пока нет
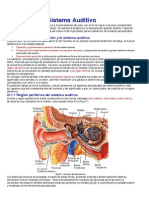
- Fisiologia Del Sistema AuditivoДокумент11 страницFisiologia Del Sistema Auditivon_almagroОценок пока нет
- Hospitalizacion InfantilДокумент6 страницHospitalizacion InfantilNayeli Del Solar HidalgoОценок пока нет
- Exposicion CarbohidratosДокумент13 страницExposicion CarbohidratosLisandra Bastidas0% (1)
- Informe 6 Resultados Laboratorio Metabolismo Fermentativo Bioquimica IiДокумент6 страницInforme 6 Resultados Laboratorio Metabolismo Fermentativo Bioquimica IiMONROY RAMIREZ ANDRES FELIPEОценок пока нет
- Hidratos de CarbonoДокумент4 страницыHidratos de CarbonoRodrigo Fernando Vera CarbajalОценок пока нет
- 10) Glicolisis PDFДокумент19 страниц10) Glicolisis PDFsigrid96Оценок пока нет
- CarbohidratosДокумент56 страницCarbohidratosCarla Yelixa Garnica MenecesОценок пока нет
- Producción de Ácido SuccínicoДокумент24 страницыProducción de Ácido SuccínicoDavid Ramírez100% (1)
- Analisis SensorialДокумент9 страницAnalisis SensorialSebas GtОценок пока нет
- Practica 3Документ3 страницыPractica 3Giselle Escobedo CortijoОценок пока нет
- La CeculosaДокумент8 страницLa CeculosaHuarocc Suyuri BryanОценок пока нет
- Bioquimica - CarbohidratosДокумент34 страницыBioquimica - CarbohidratoskarenОценок пока нет
- Expo IntoleranciaДокумент5 страницExpo IntoleranciaAndreaDavilaОценок пока нет
- Alimentacion Preescolar y EscolarДокумент24 страницыAlimentacion Preescolar y EscolarYeleОценок пока нет
- Serie Didáctica Qca de La VidaДокумент104 страницыSerie Didáctica Qca de La VidaCarlos Alfredo LópezОценок пока нет
- 04 Temario Olimpiadas (Química) 12-13Документ10 страниц04 Temario Olimpiadas (Química) 12-13Hector Daniel RinconОценок пока нет
- Glúcidos PDFДокумент17 страницGlúcidos PDFmatzalenОценок пока нет
- Carbos Al Estilo MilosДокумент8 страницCarbos Al Estilo MilosRandy88Оценок пока нет
- Guía 2do Parcial PrincipiosДокумент8 страницGuía 2do Parcial PrincipiosMayela Chacon de la CerdaОценок пока нет
- Caña de Azucar Unidad IДокумент40 страницCaña de Azucar Unidad IAna Rosa Lopez PerezОценок пока нет
- PRÁCTICA 8. Reacciones de Carbohidratos.Документ7 страницPRÁCTICA 8. Reacciones de Carbohidratos.Gloria Janet TlapechcoОценок пока нет
- Geo Arg - Manual Viticultura 2023Документ34 страницыGeo Arg - Manual Viticultura 2023Ibette NorttonОценок пока нет
- CarbohidratosДокумент47 страницCarbohidratosLina marcela unriza garciaОценок пока нет
- Combinados Primeras Asesorias 1Документ35 страницCombinados Primeras Asesorias 1Ashley TarazonaОценок пока нет
- Análisis Fisicoquimico y Sensorial para El Jugo de Caña de Azúcar y El BagazoДокумент7 страницAnálisis Fisicoquimico y Sensorial para El Jugo de Caña de Azúcar y El BagazoClary Huingo100% (1)
- Baño de Caramelo - Cómo Preparar Caramelo y Usarlo para Bañar FrutaДокумент7 страницBaño de Caramelo - Cómo Preparar Caramelo y Usarlo para Bañar FrutaDiego ViteriОценок пока нет
- Fabricacion de BiopolimerosДокумент17 страницFabricacion de BiopolimerosBismarck Sernaque CordovaОценок пока нет
- HeladosДокумент21 страницаHeladosDaniela Alexandra100% (1)
- Biologia 123Документ3 страницыBiologia 123dianyОценок пока нет
- Clase 2 Funciones Biológicas de Los Carbohidratos, Lípidos, Proteínas, Ácidos Nucleicos y Vitaminas.Документ37 страницClase 2 Funciones Biológicas de Los Carbohidratos, Lípidos, Proteínas, Ácidos Nucleicos y Vitaminas.Sofia Sandoval VásquezОценок пока нет
- El Azúcar InvisibleДокумент2 страницыEl Azúcar Invisiblecarmen pmОценок пока нет
- Diag 7 CciasДокумент14 страницDiag 7 CciasJessica SilvaОценок пока нет