Вам также может понравиться
- A Compilation of Ligno-Cellulose Feedstock and Related Research for Feed, Food and EnergyОт EverandA Compilation of Ligno-Cellulose Feedstock and Related Research for Feed, Food and EnergyОценок пока нет
- 2768 6698 27 5 140Документ13 страниц2768 6698 27 5 140Laís CarvalhoОценок пока нет
- Biofortification and Phytoremediation: Fang-Jie Zhao and Steve P McgrathДокумент8 страницBiofortification and Phytoremediation: Fang-Jie Zhao and Steve P McgrathfesooОценок пока нет
- Ácido Fítico y Cereales Integrales para La Controversia SanitariaДокумент8 страницÁcido Fítico y Cereales Integrales para La Controversia SanitariaCatiuscia BarrilliОценок пока нет
- Phytase For Food Application: Food Technology and Biotechnology November 2005Документ17 страницPhytase For Food Application: Food Technology and Biotechnology November 2005neharockОценок пока нет
- 29IJEAB 10820207 BiofortificationДокумент13 страниц29IJEAB 10820207 Biofortificationshwetabhattarai28Оценок пока нет
- Research Paper 5Документ9 страницResearch Paper 5Varsha RaniОценок пока нет
- Review Article: Trace Minerals and Livestock: Not Too Much Not Too LittleДокумент19 страницReview Article: Trace Minerals and Livestock: Not Too Much Not Too LittleRAJ KUMARОценок пока нет
- Biofortification Strategies for Improving Micronutrient DeficiencyДокумент16 страницBiofortification Strategies for Improving Micronutrient DeficiencyAMI PETОценок пока нет
- Balancing Safety and Efficacy of Lower Iron Doses in Food FortificationДокумент12 страницBalancing Safety and Efficacy of Lower Iron Doses in Food FortificationArturo EspinozaОценок пока нет
- 1-s2.0-S2773050623000381-mainДокумент7 страниц1-s2.0-S2773050623000381-mainMuhamad PermanaОценок пока нет
- Characterizing pH-Dependent Release Kinetics of Iron MicrocapsulesДокумент12 страницCharacterizing pH-Dependent Release Kinetics of Iron MicrocapsulesLimber Castro ChinoОценок пока нет
- Articulo Ciclo Del HierroДокумент12 страницArticulo Ciclo Del HierroJuliana BenavidesОценок пока нет
- Nutritional Components and Protein Quality Analysis of Genetically Modified Phytase MaizeДокумент12 страницNutritional Components and Protein Quality Analysis of Genetically Modified Phytase MaizeAJ ManurungОценок пока нет
- Zinc Biofortification of Maize (Zea Mays L.) : Status and ChallengesДокумент28 страницZinc Biofortification of Maize (Zea Mays L.) : Status and ChallengesMuhammad Amir MaqboolОценок пока нет
- bIOFORTIFICACION CakmakДокумент36 страницbIOFORTIFICACION CakmakVíctor CriolloОценок пока нет
- Phytate and Phytase in Fish NutritionДокумент30 страницPhytate and Phytase in Fish NutritionneharockОценок пока нет
- Potential Health Benefits and Adverse Effects Associated With Phytates 34683795Документ11 страницPotential Health Benefits and Adverse Effects Associated With Phytates 34683795Alain de TuringОценок пока нет
- Peptide-Metal Complexes Obtention and Role inДокумент21 страницаPeptide-Metal Complexes Obtention and Role inDiễm SươngОценок пока нет
- Advances in Human Milk Fortification With Added SupplementsДокумент9 страницAdvances in Human Milk Fortification With Added SupplementsInternational Journal of Innovative Science and Research TechnologyОценок пока нет
- Kandungan Unsur Mineral Seng (ZN), Bioavailabilitas Dan Biofortifikasinya Dalam Beras Liyanan, Elis Septianingrum Dan Bram KusbiantoroДокумент9 страницKandungan Unsur Mineral Seng (ZN), Bioavailabilitas Dan Biofortifikasinya Dalam Beras Liyanan, Elis Septianingrum Dan Bram KusbiantoroAhmad FauziОценок пока нет
- Plant Proteins As High-Quality Nutritional Source For Human DietДокумент15 страницPlant Proteins As High-Quality Nutritional Source For Human Dietjoannamae molagaОценок пока нет
- Gut Microbiota, Diet, and Health: Application To Livestock and Companion AnimalsДокумент4 страницыGut Microbiota, Diet, and Health: Application To Livestock and Companion Animalsabdelaziz CHELIGHOUMОценок пока нет
- In A Cup of Rice-The Golden Benefits of Genetically Modified Grains - Golden Rice Summary - Benzon, Consulta, Gata, Monreal, Nipas, OngonionДокумент3 страницыIn A Cup of Rice-The Golden Benefits of Genetically Modified Grains - Golden Rice Summary - Benzon, Consulta, Gata, Monreal, Nipas, OngonionSephyОценок пока нет
- Animal Nutrition: Daniel Brugger, Wilhelm M. WindischДокумент6 страницAnimal Nutrition: Daniel Brugger, Wilhelm M. WindischСнежанаОценок пока нет
- Kumari 2020Документ11 страницKumari 2020Ronald SJОценок пока нет
- Environmental Law-Internal Assessment IiiДокумент17 страницEnvironmental Law-Internal Assessment IiiAnwita BhattacharyyaОценок пока нет
- KGMC - 13 - 2009418 (About Chromium)Документ11 страницKGMC - 13 - 2009418 (About Chromium)AJ ManurungОценок пока нет
- Food Chemistry: Kaleab Baye, Jean-Pierre Guyot, Christèle Icard-Vernière, Isabelle Rochette, Claire Mouquet-RivierДокумент8 страницFood Chemistry: Kaleab Baye, Jean-Pierre Guyot, Christèle Icard-Vernière, Isabelle Rochette, Claire Mouquet-RivierYoseph AssefaОценок пока нет
- Mineral FoodsДокумент12 страницMineral FoodsARIОценок пока нет
- Adesogan2019 - Symposium Review Technologies For Improving Fiber UtilizationДокумент30 страницAdesogan2019 - Symposium Review Technologies For Improving Fiber Utilizationabdelaziz CHELIGHOUMОценок пока нет
- 2013 Article 6Документ10 страниц2013 Article 6celestinogarciaОценок пока нет
- Carotenoids: Advances in Nutrition July 2013Документ4 страницыCarotenoids: Advances in Nutrition July 2013Toàn MinhОценок пока нет
- RT Vol. 10, No. 3 Grain of TruthДокумент1 страницаRT Vol. 10, No. 3 Grain of TruthRice TodayОценок пока нет
- Mycoprotein As A Meat Substitute Production Functional Properties and CurrentДокумент23 страницыMycoprotein As A Meat Substitute Production Functional Properties and CurrentBabuskin SrinivasanОценок пока нет
- Trends in food science & technology review: Mineral chelating peptidesДокумент14 страницTrends in food science & technology review: Mineral chelating peptidesDiễm SươngОценок пока нет
- Scientia HorticulturaeДокумент8 страницScientia HorticulturaeivoneОценок пока нет
- Molecules: Enrichment of Zinc and Iron Micronutrients in Lentil (Lens Culinaris Medik.) Through BiofortificationДокумент12 страницMolecules: Enrichment of Zinc and Iron Micronutrients in Lentil (Lens Culinaris Medik.) Through BiofortificationMohammedОценок пока нет
- Traditional Food-Processing and Preparation Practices To Enhance The Bioavailability of Micronutrients in Plant-Based DietsДокумент4 страницыTraditional Food-Processing and Preparation Practices To Enhance The Bioavailability of Micronutrients in Plant-Based DietsJherson Gustavo SánchezОценок пока нет
- Artigo Aquatic Animal Nutrition and Feeding For Sustainable Aquaculture ProductionДокумент8 страницArtigo Aquatic Animal Nutrition and Feeding For Sustainable Aquaculture ProductionGuilherme HeluyОценок пока нет
- 1 s2.0 S0308814622001972 MainДокумент15 страниц1 s2.0 S0308814622001972 Mainlani wardaniОценок пока нет
- Plant Breeding To Control Zinc Deficiency in India How Cost Effective Is BiofortificationДокумент10 страницPlant Breeding To Control Zinc Deficiency in India How Cost Effective Is BiofortificationJohar ShergillОценок пока нет
- Qiu, 2020 PDFДокумент9 страницQiu, 2020 PDFHeraldo OliveiraОценок пока нет
- Industrial Applications of Marine Carbohydrates: Prasad N. Sudha, S. Aisverya, R. Nithya, K. VijayalakshmiДокумент37 страницIndustrial Applications of Marine Carbohydrates: Prasad N. Sudha, S. Aisverya, R. Nithya, K. VijayalakshmiMD. SAKIB MUBALLIGОценок пока нет
- Potential Use of Silver Nanoparticles As An Additive in Animal FeedingДокумент11 страницPotential Use of Silver Nanoparticles As An Additive in Animal FeedingSinfoJrОценок пока нет
- Society For in Vitro BiologyДокумент18 страницSociety For in Vitro Biologyanurag srivastavaОценок пока нет
- Abdullahi Muhammad Idris ProjectДокумент28 страницAbdullahi Muhammad Idris ProjectCordelia benardОценок пока нет
- Fermentation 07 00063Документ13 страницFermentation 07 00063mastersingsongОценок пока нет
- Effect of Mixed Starters On Proteolysis and Formation of Biogenic Amines in Dry Fermented Mutton SausagesДокумент15 страницEffect of Mixed Starters On Proteolysis and Formation of Biogenic Amines in Dry Fermented Mutton SausagesRoberto DegenhardtОценок пока нет
- Improving Bioavailability of Nutrients in Plant Fo PDFДокумент10 страницImproving Bioavailability of Nutrients in Plant Fo PDFTemesgen DeguОценок пока нет
- Bio 417 L14 2022Документ8 страницBio 417 L14 2022Florence Gaone GaongalelweОценок пока нет
- Fnut 09 809058Документ19 страницFnut 09 809058Stephanie Campos MalikОценок пока нет
- Paper 1Документ6 страницPaper 1neharockОценок пока нет
- Bacteria As An Alternate Biofactory For Carotenoid Production A Review of Its App, Opportu and Challenges2020Документ13 страницBacteria As An Alternate Biofactory For Carotenoid Production A Review of Its App, Opportu and Challenges2020InesОценок пока нет
- Ciência e Tecnologia de Alimentos 0101-2061: IssnДокумент9 страницCiência e Tecnologia de Alimentos 0101-2061: IssnJoseph LuiОценок пока нет
- The Importance of Minerals in Human NutritionДокумент14 страницThe Importance of Minerals in Human NutritionMargarita SosaОценок пока нет
- Kusmayadi 2021Документ9 страницKusmayadi 2021celestinogarciaОценок пока нет
- Faba Bean starch content and amino acid profilingДокумент27 страницFaba Bean starch content and amino acid profilingalfie19820223Оценок пока нет
- Fpls 12 682842Документ20 страницFpls 12 682842Nama RajasekharОценок пока нет
- P3. Membrane TransportДокумент6 страницP3. Membrane TransportCrisОценок пока нет
- CIP Annual Report 2007Документ96 страницCIP Annual Report 2007cip-libraryОценок пока нет
- 1 2 Receptor Tyrosine Kinases 2021Документ44 страницы1 2 Receptor Tyrosine Kinases 2021Bilakovics NoemiОценок пока нет
- Contoh Soalan Esei Dan Cara JawabДокумент51 страницаContoh Soalan Esei Dan Cara JawabRohayati Mamat88% (8)
- Karakterisasi Koleksi Plasma Nutfah Tomat Lokal Dan IntroduksiДокумент7 страницKarakterisasi Koleksi Plasma Nutfah Tomat Lokal Dan IntroduksiAlexanderОценок пока нет
- Body Condition Scoring BCS FinalДокумент12 страницBody Condition Scoring BCS FinalHamza HarbОценок пока нет
- Revised TERM PAPERДокумент19 страницRevised TERM PAPERVikal RajputОценок пока нет
- Vol2 Final 19092016 PDFДокумент649 страницVol2 Final 19092016 PDFshankarОценок пока нет
- Synthesis and Characterization of EDTA ComplexesДокумент7 страницSynthesis and Characterization of EDTA ComplexesLucas PereiraОценок пока нет
- Extraocular Muscles Anatomy PDFДокумент2 страницыExtraocular Muscles Anatomy PDFCherylОценок пока нет
- AP Bio Lab 1Документ5 страницAP Bio Lab 1Kiran ShilaОценок пока нет
- Malting Process PDFДокумент9 страницMalting Process PDFRay TaipeОценок пока нет
- Quiz - Mosndarja e Kromozomeve Gjate MejozwsДокумент5 страницQuiz - Mosndarja e Kromozomeve Gjate MejozwsleartaОценок пока нет
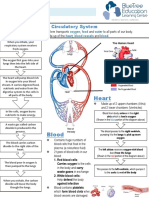
- Circulatory System Notes SLДокумент1 страницаCirculatory System Notes SLRathna PalaniappanОценок пока нет
- Case Study On Dengue FeverДокумент76 страницCase Study On Dengue FeverMary Rose Silva Gargar0% (1)
- The Digestive System Challenges and OpportunitiesДокумент9 страницThe Digestive System Challenges and OpportunitiesPhelipe PessanhaОценок пока нет
- Competitive Intelligence AcceraДокумент29 страницCompetitive Intelligence AcceranarenebiowebОценок пока нет
- Table Summary of AnatomyДокумент6 страницTable Summary of AnatomyVictor De JesusОценок пока нет
- Application of Edible Films and Coatings On MeatsДокумент2 страницыApplication of Edible Films and Coatings On MeatsAnasZeidОценок пока нет
- Comprehension QuestionДокумент2 страницыComprehension QuestionAhmad Safuan Bin Mohd Sukri -Оценок пока нет
- Neuroanatomy MCQДокумент7 страницNeuroanatomy MCQi can always make u smile :D78% (9)
- Chapter 15 - TeratogenesisДокумент29 страницChapter 15 - TeratogenesisMaula AhmadОценок пока нет
- Effect of Gender On Short Term Memory FinishedДокумент27 страницEffect of Gender On Short Term Memory Finishedhafsah286Оценок пока нет
- Biological Classification Class 11 Notes BiologyДокумент6 страницBiological Classification Class 11 Notes BiologyVyjayanthiОценок пока нет
- Quantitative Analysis of The Relative Effectiveness of 3 Iliotibial Band StretchesДокумент4 страницыQuantitative Analysis of The Relative Effectiveness of 3 Iliotibial Band Stretchessalthill01Оценок пока нет
- 1 Bio400 Sept 2018 Lecture 1 Introduction Sept 3Документ89 страниц1 Bio400 Sept 2018 Lecture 1 Introduction Sept 3The seriОценок пока нет
- P - Science - 5 - Worksheets - Unit 1Документ18 страницP - Science - 5 - Worksheets - Unit 1Jaishree Rathi80% (5)
- Physiology - Last Minute Revision Points: DR - SreetejaДокумент25 страницPhysiology - Last Minute Revision Points: DR - SreetejaBanu KubendiranОценок пока нет
- Science Reviewer - FinaДокумент3 страницыScience Reviewer - FinaAmamore Lorenzana PlazaОценок пока нет
- Lipid PresentationДокумент20 страницLipid PresentationFrances Anne OrbinoОценок пока нет