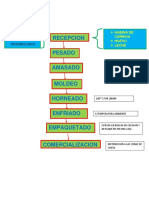
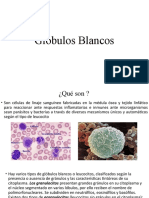
Вам также может понравиться
- 3.01.02 Enfermedad AujeszkyДокумент19 страниц3.01.02 Enfermedad AujeszkyDaniel JaramilloОценок пока нет
- 3.08.02 Artritis-Encefalitis Caprina Maedi VisnaДокумент10 страниц3.08.02 Artritis-Encefalitis Caprina Maedi Visnatecnicos.tolima.huilaОценок пока нет
- 2.07.10. Adenocarcinoma Pulmonar OvinoДокумент5 страниц2.07.10. Adenocarcinoma Pulmonar OvinoJavier Manrique SalamancaОценок пока нет
- 3.01.17 RabiesДокумент38 страниц3.01.17 RabiesChristhell SanchezОценок пока нет
- 3 Síndrome Reproductivo y Respiratorio Porcino IIДокумент6 страниц3 Síndrome Reproductivo y Respiratorio Porcino IIAxoncomunicacionОценок пока нет
- 3.04.02 - Babesiosis BovinaДокумент18 страниц3.04.02 - Babesiosis BovinaMardex CoCОценок пока нет
- 3.08.02 Rinitis Atrófica Porcina PDFДокумент12 страниц3.08.02 Rinitis Atrófica Porcina PDFCruz Eliud MachadoОценок пока нет
- Nipah EnfermedadesДокумент11 страницNipah EnfermedadesJAYNARA FRANCOОценок пока нет
- 3.06.01 Peste EquinaДокумент17 страниц3.06.01 Peste EquinaDaniel JaramilloОценок пока нет
- E Schmallenberg VirusДокумент5 страницE Schmallenberg Viruscasandra.g.momaxОценок пока нет
- 3.01.01 Carbunco BacteridianoДокумент15 страниц3.01.01 Carbunco BacteridianoMelisa ParadaОценок пока нет
- 3.01.08 - Fiebre AftosaДокумент37 страниц3.01.08 - Fiebre AftosaDaniel JaramilloОценок пока нет
- 3.04.04 Campilobact BovinaДокумент13 страниц3.04.04 Campilobact BovinaValeria NavarroОценок пока нет
- Mesonero Escuredo Et Al - 2010 - Parvovirus Porcino Parte IIДокумент5 страницMesonero Escuredo Et Al - 2010 - Parvovirus Porcino Parte IITatianaОценок пока нет
- ADENOMATOSIS PULMONAR OVINA (Jaagsiekte)Документ12 страницADENOMATOSIS PULMONAR OVINA (Jaagsiekte)Darwin Antezana De la RosaОценок пока нет
- Epidemiología y Diagnóstico de IbrДокумент3 страницыEpidemiología y Diagnóstico de Ibrjorge urbinaОценок пока нет
- Seminario Diagnostico VirológicoДокумент15 страницSeminario Diagnostico VirológicoPetter VahlisОценок пока нет
- Cuestionario-Diagnostico ViralДокумент3 страницыCuestionario-Diagnostico ViralRoberta VianaОценок пока нет
- Grupo C4 Sesión de Aprendizaje N°13 - PCR para VPH y Pruebas para CMVДокумент9 страницGrupo C4 Sesión de Aprendizaje N°13 - PCR para VPH y Pruebas para CMVLeyla Barnard LagunaОценок пока нет
- Tema 8.5. Infecciones Respiratorias - ParvoviridaeДокумент11 страницTema 8.5. Infecciones Respiratorias - ParvoviridaeAlexandra AliagaОценок пока нет
- 2 Doparcial 1 Ercuat 2007Документ16 страниц2 Doparcial 1 Ercuat 2007Goa joyasОценок пока нет
- Univesidad Nacional de San Antonio Abad Del Cusco: Facultad de Ciencias AgrariasДокумент18 страницUnivesidad Nacional de San Antonio Abad Del Cusco: Facultad de Ciencias AgrariasNelson Roger Saire ChoquenairaОценок пока нет
- 2 Bronquitis Infecciosa AviarДокумент17 страниц2 Bronquitis Infecciosa AviarJavi CarrilloОценок пока нет
- 3.01.12 - Leptospirosis OIEДокумент16 страниц3.01.12 - Leptospirosis OIEAnna Diaz YuberoОценок пока нет
- AstrovirusДокумент8 страницAstrovirusLucía Margarita VillaОценок пока нет
- Tecnica MolecularДокумент4 страницыTecnica MolecularCarlos reyes archundiaОценок пока нет
- TAREA Nro 11 Diagnostico de EI en Distinatas EspeciesДокумент5 страницTAREA Nro 11 Diagnostico de EI en Distinatas EspeciesAnairis ManriqueОценок пока нет
- Vigilancia de La PPAДокумент60 страницVigilancia de La PPAangelamorillopoche08Оценок пока нет
- La Técnica de ELISAДокумент6 страницLa Técnica de ELISAAndhy LoorОценок пока нет
- Ficha Tecnica DEPДокумент4 страницыFicha Tecnica DEPLaura AguileraОценок пока нет
- Parvovirosis en Caninos y Porcinos - PPTX PRESENTACIONДокумент14 страницParvovirosis en Caninos y Porcinos - PPTX PRESENTACIONFranklinCopaОценок пока нет
- 5 BPSANIDADenCerdosДокумент42 страницы5 BPSANIDADenCerdosCarlosОценок пока нет
- Maedi Visna AECДокумент2 страницыMaedi Visna AECDalotodo OnohagasnadaОценок пока нет
- Pruebas de Adn para El Virus Del Papiloma Humano. Retos en La Tamizacion Del Vph. Aurora Aznar FuenlabradaДокумент34 страницыPruebas de Adn para El Virus Del Papiloma Humano. Retos en La Tamizacion Del Vph. Aurora Aznar FuenlabradaAurora aznarОценок пока нет
- Métodos Diagnósticos Utilizados en AviculturaДокумент9 страницMétodos Diagnósticos Utilizados en AviculturaLiceth ParraОценок пока нет
- PRACTICA 16 y 17 Enfermeria 2022Документ29 страницPRACTICA 16 y 17 Enfermeria 2022Iveth Cortez CuevaОценок пока нет
- Virus Del Papiloma Humano ResumenДокумент7 страницVirus Del Papiloma Humano ResumenEduardo MiramontesОценок пока нет
- Virus IntestinalesДокумент4 страницыVirus IntestinalesFranche OportoОценок пока нет
- Introducción Metodos Diagnósticos y ParásitosДокумент33 страницыIntroducción Metodos Diagnósticos y ParásitosPatricia Fernández TapiaОценок пока нет
- Pruebas Diagnósticas para VihДокумент3 страницыPruebas Diagnósticas para VihCésar LezamaОценок пока нет
- Aplicaciones Wester BlotДокумент2 страницыAplicaciones Wester BlotNoemiMirandaMeloОценок пока нет
- Of - Rotavirus Lucia MarquezДокумент17 страницOf - Rotavirus Lucia Marquezlucia marquez lopezОценок пока нет
- Epidemiologia de ScrapieДокумент1 страницаEpidemiologia de Scrapieamaru gonzalezОценок пока нет
- Caso Clínico de Anatomía Patológica PRRS ANAPORC 181Документ6 страницCaso Clínico de Anatomía Patológica PRRS ANAPORC 181Jose Jesus Martinez NunezОценок пока нет
- F Tecnica Diarrea Epidemica Porcina-2018Документ2 страницыF Tecnica Diarrea Epidemica Porcina-2018Cristopher MuñozОценок пока нет
- Vigilancia Epidemiologica Enfermedades ZoonoticasДокумент20 страницVigilancia Epidemiologica Enfermedades ZoonoticasSamuel Timpo PauccarОценок пока нет
- 687 2564 1 PBДокумент12 страниц687 2564 1 PBLinbert Simon CallataОценок пока нет
- Paratuberculosis Es PDFДокумент6 страницParatuberculosis Es PDFpablobhaОценок пока нет
- Diarrea Viral BovinaДокумент8 страницDiarrea Viral BovinaYairis RiveraОценок пока нет
- Vacuna de La Rabia FolletoДокумент5 страницVacuna de La Rabia Folletotiroquim563Оценок пока нет
- Infecciones Viricas GastrointestinalesДокумент2 страницыInfecciones Viricas GastrointestinalesRomi OrtizОценок пока нет
- Plan de Control y Prevencion de La RabiaДокумент33 страницыPlan de Control y Prevencion de La RabiaLugan VetsОценок пока нет
- Loque EuropeaДокумент9 страницLoque EuropeaKaryme Morales JaramilloОценок пока нет
- Enfermedad de Newcastle en PoinДокумент31 страницаEnfermedad de Newcastle en PoinAlex Amaya100% (1)
- Pruebas de Infectividad en VirusДокумент15 страницPruebas de Infectividad en VirusRomulo Aycachi Inga100% (3)
- Anti Micro Bi A NosДокумент62 страницыAnti Micro Bi A NoseialcaheОценок пока нет
- 3.01.12 LeptospirosisДокумент14 страниц3.01.12 LeptospirosisJenni De La O BautistaОценок пока нет
- Virus del Nilo Occidental ¿Estamos preparados?: Una perspectiva One HealthОт EverandVirus del Nilo Occidental ¿Estamos preparados?: Una perspectiva One HealthОценок пока нет
- Técnicas para el diagnóstico de endoparásitos de importancia veterinariaОт EverandTécnicas para el diagnóstico de endoparásitos de importancia veterinariaОценок пока нет
- Ley de Endeudamiento PublicoДокумент5 страницLey de Endeudamiento Publicogreco caillahua chavez100% (1)
- CeftriaxonaДокумент6 страницCeftriaxonaEduardo Vargas Ortiz100% (1)
- Revges GP Febrerorevista2014 18Документ4 страницыRevges GP Febrerorevista2014 18justo danielОценок пока нет
- Artículos OriginalesДокумент12 страницArtículos Originalesjusto danielОценок пока нет
- Doc1 ANYДокумент1 страницаDoc1 ANYjusto danielОценок пока нет
- ApartadosДокумент12 страницApartadosjusto danielОценок пока нет
- Inta-Produccionovina IntaДокумент5 страницInta-Produccionovina Intajusto danielОценок пока нет
- El Medio Ambiente en La Proteccion de AlimentosДокумент10 страницEl Medio Ambiente en La Proteccion de Alimentosjusto danielОценок пока нет
- El Medio Ambiente en La Proteccion de AlimentosДокумент5 страницEl Medio Ambiente en La Proteccion de Alimentosjusto danielОценок пока нет
- Contenido TemáticoДокумент23 страницыContenido Temáticojusto danielОценок пока нет
- Anemia Leve Causado Por Piojos en Alpaca CorregidoДокумент10 страницAnemia Leve Causado Por Piojos en Alpaca Corregidojusto danielОценок пока нет
- Burgos 2017 Anestesia Segura en El Paciente FelinoДокумент14 страницBurgos 2017 Anestesia Segura en El Paciente FelinorockyОценок пока нет
- Artículos PrimariosДокумент11 страницArtículos Primariosjusto danielОценок пока нет
- Artículos PrimariosДокумент11 страницArtículos Primariosjusto danielОценок пока нет
- Informe CosoДокумент16 страницInforme CosoNorberto Ruiz0% (1)
- Diureticos PT EsДокумент6 страницDiureticos PT Esjusto danielОценок пока нет
- Inta-Produccionovina Inta PDFДокумент221 страницаInta-Produccionovina Inta PDFAlberto LobonesОценок пока нет
- Univ - Chile Tesis Guia Del Estudio de Mercado para La Evaluacion de ProyectosДокумент149 страницUniv - Chile Tesis Guia Del Estudio de Mercado para La Evaluacion de ProyectosANiita GAbiiela ArriiagaОценок пока нет
- El Medio Ambiente en La Proteccion de AlimentosДокумент10 страницEl Medio Ambiente en La Proteccion de Alimentosjusto danielОценок пока нет
- 625Документ142 страницы625Remy AQОценок пока нет
- Burgos 2017 Anestesia Segura en El Paciente FelinoДокумент92 страницыBurgos 2017 Anestesia Segura en El Paciente Felinojusto danielОценок пока нет
- HACCP Cereales ExtruidosДокумент92 страницыHACCP Cereales ExtruidosHans100% (2)
- El Medio Ambiente en La Proteccion de AlimentosДокумент10 страницEl Medio Ambiente en La Proteccion de Alimentosjusto danielОценок пока нет
- 1 Modelos de ControlДокумент117 страниц1 Modelos de Controlelyrueda80% (10)
- Acta ConstitucionДокумент2 страницыActa ConstitucionCesar EstradaОценок пока нет
- Formato de Minuta SAC Con Directorio Aporte BienesДокумент7 страницFormato de Minuta SAC Con Directorio Aporte Bienesjusto daniel33% (3)
- Sistema de Control InternoДокумент50 страницSistema de Control InternojockergapОценок пока нет
- Filminas - Control InternoДокумент20 страницFilminas - Control InternoMary SalcedoОценок пока нет
- Biología Re y GolgiДокумент6 страницBiología Re y GolgiNabila InsfranОценок пока нет
- Tarea Células y Componentes Solubles de La Respuesta InmuneДокумент8 страницTarea Células y Componentes Solubles de La Respuesta InmuneRaquel itzamar Salazar LópezОценок пока нет
- Guía de Biología Pedro ValdiviaДокумент28 страницGuía de Biología Pedro ValdiviaCatherine Rangel0% (1)
- Buenas Practicas de Manejo en AvesДокумент22 страницыBuenas Practicas de Manejo en AvesPaula María100% (2)
- Tema 02 LA CÉLULA Y LOS TEJIDOS. LA PIEL Y SU PATOLOGÍAДокумент23 страницыTema 02 LA CÉLULA Y LOS TEJIDOS. LA PIEL Y SU PATOLOGÍAFrauleinОценок пока нет
- MacrófagoДокумент5 страницMacrófagocieloОценок пока нет
- Introducciã"n A La Genã Tica ...............Документ32 страницыIntroducciã"n A La Genã Tica ...............Gonzalo MenendezОценок пока нет
- Organismos de Interes BiotecnologicoДокумент31 страницаOrganismos de Interes BiotecnologicoJuliet LCОценок пока нет
- 6alteracionesdehematies 110702110852 Phpapp02 PDFДокумент28 страниц6alteracionesdehematies 110702110852 Phpapp02 PDFJael Sepulveda HidalgoОценок пока нет
- Reino Monera Trabajamos en ClaseДокумент6 страницReino Monera Trabajamos en Clasewr64203Оценок пока нет
- Estructura BacterianaДокумент8 страницEstructura BacterianaJesus Rafael AstudilloОценок пока нет
- Los Cromosomas Guia 9Документ3 страницыLos Cromosomas Guia 9genovitaОценок пока нет
- Glóbulos BlancosДокумент18 страницGlóbulos BlancosRenato Benavides OlivaОценок пока нет
- Celula EucariotaДокумент6 страницCelula EucariotaMelissa Poma50% (2)
- Cromosoma 13Документ16 страницCromosoma 13api-26666279Оценок пока нет
- Morfologia Del Sistema Hematopoyetico PDFДокумент44 страницыMorfologia Del Sistema Hematopoyetico PDFBen CórdovaОценок пока нет
- Neumonia Adquirida en La Comunidad: MR Donayre Hospital Maria AuxiliadoraДокумент39 страницNeumonia Adquirida en La Comunidad: MR Donayre Hospital Maria AuxiliadoraSamantha DonayreОценок пока нет
- Ensayo Vacunación y EmbarazoДокумент10 страницEnsayo Vacunación y EmbarazoNatalia DiazОценок пока нет
- Complejo Respiratorio BovinoДокумент5 страницComplejo Respiratorio BovinoMatias Ruiz-Tagle GuerreroОценок пока нет
- KLEBSIELLAДокумент12 страницKLEBSIELLAClarita GallardoОценок пока нет
- USMLE STEP 3 Lecture Notes 2017-2018Документ4 страницыUSMLE STEP 3 Lecture Notes 2017-2018ArnoldTito100% (2)
- Cómo Eliminar Las Verrugas de La Cara - Cómo QuitarlasДокумент3 страницыCómo Eliminar Las Verrugas de La Cara - Cómo Quitarlastratamientos naturalesОценок пока нет
- Sesión #10: FagocitosisДокумент59 страницSesión #10: FagocitosisPretty LitiñaОценок пока нет
- Aminoacidos, Peptidos y ProteinasДокумент21 страницаAminoacidos, Peptidos y ProteinasFlorencia Barsi100% (2)
- Examen Final de BiologiaДокумент10 страницExamen Final de BiologiaCarlos AlmanzaОценок пока нет
- T2 Protistas Protofitos y ProtozoosДокумент23 страницыT2 Protistas Protofitos y Protozoosmiguel angel tejerina100% (1)
- Evaluación GenéticaДокумент6 страницEvaluación GenéticaCarlos Eduardo Rincon Obando0% (1)
- SerologiaДокумент14 страницSerologiaBrandon Isaid Reyna VillafuerteОценок пока нет
- Escuela Luis Oyarzún Peña Dpto. de Orientación Y Promoción de Salud Lorena Arenas Lopez - Carolina Díaz SilvaДокумент12 страницEscuela Luis Oyarzún Peña Dpto. de Orientación Y Promoción de Salud Lorena Arenas Lopez - Carolina Díaz SilvaLorena Arenas LopezОценок пока нет
- DominiosДокумент15 страницDominiosIvan LosadaОценок пока нет