Вам также может понравиться
- 9037 PDFДокумент6 страниц9037 PDFpdfОценок пока нет
- Moisture - Diffusion - in - Epoxy - SystemsДокумент13 страницMoisture - Diffusion - in - Epoxy - SystemsVansala GanesanОценок пока нет
- Art 681Документ10 страницArt 681Lungelo DlaminiОценок пока нет
- Journal Homepage: - : IntroductionДокумент5 страницJournal Homepage: - : IntroductionIJAR JOURNALОценок пока нет
- Pre041908 2009Документ4 страницыPre041908 2009tnbtnbtnbОценок пока нет
- Protein Adhesion to Biomaterial Surfaces Impacted by Surface WettabilityДокумент11 страницProtein Adhesion to Biomaterial Surfaces Impacted by Surface WettabilityKhay NochefrancaОценок пока нет
- AdsorcionДокумент9 страницAdsorcionMary FallОценок пока нет
- 1 s2.0 S0376738804000523 MainДокумент9 страниц1 s2.0 S0376738804000523 MainchetanОценок пока нет
- PhysRevFluids 3 123102Документ21 страницаPhysRevFluids 3 123102hifan756Оценок пока нет
- How pH Impacts Protein Solvent Flux in Stirred UltrafiltrationДокумент8 страницHow pH Impacts Protein Solvent Flux in Stirred UltrafiltrationRoxana ElenaОценок пока нет
- Confocal 04 PDFДокумент7 страницConfocal 04 PDFAl-somuda' Ali Al-smmaniОценок пока нет
- 1 EmmanuelPowerAfricaIEEEДокумент7 страниц1 EmmanuelPowerAfricaIEEEمحمد أشرفОценок пока нет
- Protein AnalysisДокумент17 страницProtein AnalysisAfrina アデリナОценок пока нет
- J. H. Booske Et Al - Microwave Ponderomotive Forces in Solid-State Ionic PlasmasДокумент7 страницJ. H. Booske Et Al - Microwave Ponderomotive Forces in Solid-State Ionic PlasmasDublin000Оценок пока нет
- TMP 70 D3Документ20 страницTMP 70 D3FrontiersОценок пока нет
- Electrochemical Behaviour of HAEMOGLOGIN 2008Документ6 страницElectrochemical Behaviour of HAEMOGLOGIN 2008ericОценок пока нет
- Physica B: M.H. Buraidah, L.P. Teo, S.R. Majid, A.K. ArofДокумент7 страницPhysica B: M.H. Buraidah, L.P. Teo, S.R. Majid, A.K. ArofArindam SenОценок пока нет
- Chen, H., Langer, R. & Edwards, D. A. A Film Tension Theory of Phagocytosis.Документ1 страницаChen, H., Langer, R. & Edwards, D. A. A Film Tension Theory of Phagocytosis.DenisОценок пока нет
- Freger 2008 Polymer Dowex50Документ12 страницFreger 2008 Polymer Dowex50Axel EttoriОценок пока нет
- Flicking Er 1999Документ23 страницыFlicking Er 1999Margaret RogersОценок пока нет
- Biochemistry CH 3Документ3 страницыBiochemistry CH 3Ann Ross FernandezОценок пока нет
- Template Technical ReportДокумент6 страницTemplate Technical ReportZarith IhsanОценок пока нет
- Effects of The Antimalarial Drug Primaquine On The Dynamic Structure of Lipid Model MembranesДокумент10 страницEffects of The Antimalarial Drug Primaquine On The Dynamic Structure of Lipid Model MembranesFadi Simon de Souza MagalhãesОценок пока нет
- Huang 2008Документ12 страницHuang 2008Hamid ChenaraniОценок пока нет
- The Development of Polycarbophil As A Bioadhesive MaterialДокумент10 страницThe Development of Polycarbophil As A Bioadhesive MaterialCesar Rodolfo Angulo DelgadoОценок пока нет
- 2012 Dep of Conductivity On Charge Density and Echem Pot in Polymer SC W Ionic LiquidsДокумент10 страниц2012 Dep of Conductivity On Charge Density and Echem Pot in Polymer SC W Ionic LiquidsChris SmithОценок пока нет
- NafionДокумент9 страницNafionAlejandro BedollaОценок пока нет
- Mechanisms For The Ultrasonic Enhancement of Dairy Whey UltrafiltrationДокумент9 страницMechanisms For The Ultrasonic Enhancement of Dairy Whey UltrafiltrationTimmyNguyenОценок пока нет
- Bacteria SwimmingДокумент4 страницыBacteria Swimmingparis.hiltonОценок пока нет
- Electrical Properties of Tissues and Cell Suspensions - Schwan2Документ2 страницыElectrical Properties of Tissues and Cell Suspensions - Schwan2Douglas DutraОценок пока нет
- Potential For Modulation of The Hydrophobic Effect Inside ChaperoninsДокумент9 страницPotential For Modulation of The Hydrophobic Effect Inside Chaperoninsmasteranza5936Оценок пока нет
- Electrical Properties of Tissues and Cell Suspensions: Mechanisms and ModelsДокумент2 страницыElectrical Properties of Tissues and Cell Suspensions: Mechanisms and Modelspro.gig9033Оценок пока нет
- Concentration and Desalination of Protein Solutions by UltrafiltrationДокумент11 страницConcentration and Desalination of Protein Solutions by UltrafiltrationClarence AG YueОценок пока нет
- 216 2016 Article 9464Документ9 страниц216 2016 Article 9464restiОценок пока нет
- ROLE OF POLYMER MATRIXДокумент7 страницROLE OF POLYMER MATRIXhanayuki007Оценок пока нет
- (The Plenum Chemical Engineering Series) Y. T. Shah, A. B. Pandit, V. S. Moholkar (Auth.) - Cavitation Reaction Engineering-Springer US (1999) PDFДокумент362 страницы(The Plenum Chemical Engineering Series) Y. T. Shah, A. B. Pandit, V. S. Moholkar (Auth.) - Cavitation Reaction Engineering-Springer US (1999) PDFjuan camilo bedoya cardonaОценок пока нет
- 2019 - AEM - PCE11.o-IDTBR-MW and Miscibility Correlating With Device Performance and MorphologyДокумент14 страниц2019 - AEM - PCE11.o-IDTBR-MW and Miscibility Correlating With Device Performance and MorphologyBilal NaveedОценок пока нет
- Jacs 7b09508Документ9 страницJacs 7b09508elderwanОценок пока нет
- Gurt Flip Flop Jpcb07Документ6 страницGurt Flip Flop Jpcb07Dinok DiblongОценок пока нет
- Can e Guner 2012Документ12 страницCan e Guner 2012Vithória Carolyna Trindade Dos SantosОценок пока нет
- Relationship Between Lemna Growth and pHДокумент6 страницRelationship Between Lemna Growth and pHMark CwmОценок пока нет
- Water Segregation Attic Greek: Hydrophobic InteractionДокумент11 страницWater Segregation Attic Greek: Hydrophobic InteractionHemshree PrasadОценок пока нет
- Mechanisms of Proton Conductance in Polymer Electrolyte MembranesДокумент17 страницMechanisms of Proton Conductance in Polymer Electrolyte MembranesDeva RajОценок пока нет
- Membrane Potentials in Living Systems, Tools To Measure: Biological BackgroundДокумент10 страницMembrane Potentials in Living Systems, Tools To Measure: Biological BackgroundazzaassОценок пока нет
- Electric Fields and Vortex Mixing Improve Ultrafiltration RatesДокумент10 страницElectric Fields and Vortex Mixing Improve Ultrafiltration RatesJaime JuradoОценок пока нет
- 2.2.47. Capillary ElectrophoresisДокумент6 страниц2.2.47. Capillary ElectrophoresisLuizaОценок пока нет
- Bowen Calvo Steps1995Документ14 страницBowen Calvo Steps1995Ilyes JedidiОценок пока нет
- Greenhoe Et Al-2016-Journal of Polymer Science Part B - Polymer Physics JournalwebsiteДокумент7 страницGreenhoe Et Al-2016-Journal of Polymer Science Part B - Polymer Physics JournalwebsiteMoad BarbariОценок пока нет
- UntitledДокумент30 страницUntitledGerardo LozanoОценок пока нет
- Polymers: Surface Chemistry of PolymersДокумент4 страницыPolymers: Surface Chemistry of PolymersMOHAMED DiriyeОценок пока нет
- Review Partitioning in Aqueous Two-Phase Systems: Recent ResultsДокумент18 страницReview Partitioning in Aqueous Two-Phase Systems: Recent ResultsAmanda E. SalesОценок пока нет
- tmp3539 TMPДокумент10 страницtmp3539 TMPFrontiersОценок пока нет
- Protein Nanoparticle InteractionsДокумент8 страницProtein Nanoparticle InteractionsRadu AndreiОценок пока нет
- Analysis of Human Histone H4 by Capillary Electrophoresis in A Pullulan-Coated Capillary, LC-ESI-MS and Maldi-Tof-MsДокумент8 страницAnalysis of Human Histone H4 by Capillary Electrophoresis in A Pullulan-Coated Capillary, LC-ESI-MS and Maldi-Tof-MsVic PastorОценок пока нет
- Enzymatic Hollow Fiber Membrane Bioreactor For Penicilin HydrolysisДокумент7 страницEnzymatic Hollow Fiber Membrane Bioreactor For Penicilin HydrolysisTravis ReillyОценок пока нет
- Biology 1002B OutcomesДокумент6 страницBiology 1002B OutcomesJenny JooОценок пока нет
- Articulo, Soludilu PDFДокумент2 страницыArticulo, Soludilu PDFJose Ruben CarrilloОценок пока нет
- L9. Dialysis. ElectrophoresisДокумент5 страницL9. Dialysis. ElectrophoresisanaОценок пока нет
- Bioreactor Modeling: Interactions between Hydrodynamics and BiologyОт EverandBioreactor Modeling: Interactions between Hydrodynamics and BiologyОценок пока нет
- Filtrarea TangentialaДокумент163 страницыFiltrarea TangentialaRoxana ElenaОценок пока нет
- MembraneModuleDesignandDynamicShear InducedTechniques PDFДокумент58 страницMembraneModuleDesignandDynamicShear InducedTechniques PDFRoxana ElenaОценок пока нет
- MembraneModuleDesignandDynamicShear InducedTechniques PDFДокумент58 страницMembraneModuleDesignandDynamicShear InducedTechniques PDFRoxana ElenaОценок пока нет
- Astm c1036Документ8 страницAstm c1036the lookОценок пока нет
- Ultrafiltration: Boon or Bane?: R, Vol 51, NR 2, 2009, P 613-625Документ13 страницUltrafiltration: Boon or Bane?: R, Vol 51, NR 2, 2009, P 613-625Roxana ElenaОценок пока нет
- 2011 - Concentration and Purification of Whey Proteins by UltrafiltrationДокумент6 страниц2011 - Concentration and Purification of Whey Proteins by UltrafiltrationRoxana ElenaОценок пока нет
- 2 PDFДокумент8 страниц2 PDFRoxana ElenaОценок пока нет
- Whole PDFДокумент279 страницWhole PDFRoxana ElenaОценок пока нет
- The Effect of Protein Protein and Protei PDFДокумент12 страницThe Effect of Protein Protein and Protei PDFRoxana ElenaОценок пока нет
- Journal of Water Process Engineering: SciencedirectДокумент8 страницJournal of Water Process Engineering: SciencedirectRoxana ElenaОценок пока нет
- Ultrafiltration: Boon or Bane?: R, Vol 51, NR 2, 2009, P 613-625Документ13 страницUltrafiltration: Boon or Bane?: R, Vol 51, NR 2, 2009, P 613-625Roxana ElenaОценок пока нет
- Ultrafiltration: Boon or Bane?: R, Vol 51, NR 2, 2009, P 613-625Документ13 страницUltrafiltration: Boon or Bane?: R, Vol 51, NR 2, 2009, P 613-625Roxana ElenaОценок пока нет
- High Performance Dialysis GuideДокумент28 страницHigh Performance Dialysis GuideRoxana ElenaОценок пока нет
- DetectorДокумент276 страницDetectorRoxana ElenaОценок пока нет
- Dupa Ce Ne-Am Indragostit - Todd Anna - PDДокумент18 страницDupa Ce Ne-Am Indragostit - Todd Anna - PDRoxana ElenaОценок пока нет
- WebДокумент67 страницWebRoxana ElenaОценок пока нет
- Chemical Engineering Science: Mylène Wang, Sourav Mondal, Ian M. GriffithsДокумент8 страницChemical Engineering Science: Mylène Wang, Sourav Mondal, Ian M. GriffithsRoxana ElenaОценок пока нет
- 1602 07467Документ54 страницы1602 07467Roxana ElenaОценок пока нет
- Study On The Stabilisation of Collagen With Vegetable Tannins in The Presence of Acrylic PolymerДокумент7 страницStudy On The Stabilisation of Collagen With Vegetable Tannins in The Presence of Acrylic PolymerRoxana ElenaОценок пока нет
- 02 WholeДокумент242 страницы02 WholeRoxana ElenaОценок пока нет
- Characteristics and Uses of CollagenДокумент6 страницCharacteristics and Uses of CollagenRoxana ElenaОценок пока нет
- FBTADIRCROSSENДокумент10 страницFBTADIRCROSSENRoxana ElenaОценок пока нет
- How pH Impacts Protein Solvent Flux in Stirred UltrafiltrationДокумент8 страницHow pH Impacts Protein Solvent Flux in Stirred UltrafiltrationRoxana ElenaОценок пока нет
- Study On The Stabilisation of Collagen With Vegetable Tannins in The Presence of Acrylic PolymerДокумент7 страницStudy On The Stabilisation of Collagen With Vegetable Tannins in The Presence of Acrylic PolymerRoxana ElenaОценок пока нет
- Us 5490937Документ25 страницUs 5490937Roxana ElenaОценок пока нет
- Biodegradability and Cell-Mediated Contraction of Porous Collagen Scaffolds: The Effect of Lysine As A Novel Crosslinking BridgeДокумент9 страницBiodegradability and Cell-Mediated Contraction of Porous Collagen Scaffolds: The Effect of Lysine As A Novel Crosslinking BridgeRoxana ElenaОценок пока нет
- Dialysis Tubing Prep and StorageДокумент1 страницаDialysis Tubing Prep and StorageRoxana ElenaОценок пока нет
- CollagenДокумент2 страницыCollagenRoxana ElenaОценок пока нет
- 40697Документ48 страниц40697Roxana ElenaОценок пока нет
- Forex - Nnet Vs RegДокумент6 страницForex - Nnet Vs RegAnshik BansalОценок пока нет
- Adobe After Effects CS3 Keyboard Shortcuts GuideДокумент14 страницAdobe After Effects CS3 Keyboard Shortcuts GuideBrandon Sirota100% (1)
- MP Lab3Документ4 страницыMP Lab3kawish420Оценок пока нет
- FOUNDATION REPAIR AND REGROUT FOR BODYMAKER AT CARNDAUD METALBOX SINGAPOREДокумент15 страницFOUNDATION REPAIR AND REGROUT FOR BODYMAKER AT CARNDAUD METALBOX SINGAPORETrúc NguyễnОценок пока нет
- VHDL ExperimentsДокумент55 страницVHDL Experimentssandeepsingh93Оценок пока нет
- Air Suspension Benefits Over Steel SpringsДокумент3 страницыAir Suspension Benefits Over Steel SpringsBejoy G NairОценок пока нет
- TXP TrainingДокумент88 страницTXP Trainingsina20795100% (1)
- Math Let Reviewer 44 Analytic GeometryДокумент3 страницыMath Let Reviewer 44 Analytic Geometryjohn johnОценок пока нет
- NX Advanced Simulation坐标系Документ12 страницNX Advanced Simulation坐标系jingyong123Оценок пока нет
- E8244 Ahu Catalogue 2014 PDFДокумент174 страницыE8244 Ahu Catalogue 2014 PDFmurugesanОценок пока нет
- SSC JE Mechanical Study Material Strength of Materials PDFДокумент13 страницSSC JE Mechanical Study Material Strength of Materials PDFSameer ShahОценок пока нет
- The 2009 ACM ASIA Programming Contest Dhaka Site: Hosted by North South University Dhaka, BangladeshДокумент21 страницаThe 2009 ACM ASIA Programming Contest Dhaka Site: Hosted by North South University Dhaka, BangladeshPhạm Hữu Thạnh ĐạtОценок пока нет
- SM Maintenance Instructions: Author: Lars Rydén, Konecranes AB, SwedenДокумент132 страницыSM Maintenance Instructions: Author: Lars Rydén, Konecranes AB, SwedenDan VekasiОценок пока нет
- Queue PPTДокумент19 страницQueue PPTharshvardhanshinde.2504Оценок пока нет
- Development and Validation of UV Spectrophotometric Method For The Estimation of Curcumin in Bulk Drug and Pharmaceutical Dosage FormsДокумент6 страницDevelopment and Validation of UV Spectrophotometric Method For The Estimation of Curcumin in Bulk Drug and Pharmaceutical Dosage FormswiracanaОценок пока нет
- What Is Radar and Its FunctionДокумент3 страницыWhat Is Radar and Its FunctionJean Maya DiscayaОценок пока нет
- Week 2Документ11 страницWeek 2KISHAОценок пока нет
- Lecture Antenna MiniaturizationДокумент34 страницыLecture Antenna MiniaturizationJuhi GargОценок пока нет
- Tensile Properties of Ground Coffee Waste Reinforced Polyethylene CompositeДокумент4 страницыTensile Properties of Ground Coffee Waste Reinforced Polyethylene CompositemhmmdzulvaОценок пока нет
- POWER Nav1 - MIDTERM - Topic 1 & 2Документ31 страницаPOWER Nav1 - MIDTERM - Topic 1 & 2CLIJOHN PABLO FORD100% (1)
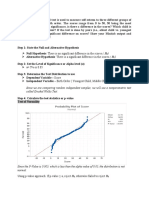
- Step 1: State The Null and Alternative HypothesisДокумент3 страницыStep 1: State The Null and Alternative HypothesisChristine Joyce BascoОценок пока нет
- Dev - Mag - 09Документ26 страницDev - Mag - 09georgpiorczynskiОценок пока нет
- Lecture 2 - Kinematics Fundamentals - Part AДокумент30 страницLecture 2 - Kinematics Fundamentals - Part ASuaid Tariq BalghariОценок пока нет
- Business Calculus NotesДокумент38 страницBusiness Calculus NotesTom KowalskiОценок пока нет
- Qualcomm Snapdragon 660 Mobile Platform Product BriefДокумент2 страницыQualcomm Snapdragon 660 Mobile Platform Product BriefTuxInfoОценок пока нет
- SD Revenue Account Determination SetupДокумент10 страницSD Revenue Account Determination SetupkarthikbjОценок пока нет
- Equipment DetailsДокумент10 страницEquipment Detailsimranjani.skОценок пока нет
- 10th Geometrical ConstructionДокумент2 страницы10th Geometrical Constructionapi-242227794100% (1)
- Barium Strontium TitanateДокумент15 страницBarium Strontium Titanatekanita_jawwОценок пока нет
- Flow MeterДокумент9 страницFlow MeterShin BoОценок пока нет