Вам также может понравиться
- Prostatitis AgudaДокумент6 страницProstatitis AgudaEliseo CMОценок пока нет
- C0v1d M4y0Документ8 страницC0v1d M4y0Eliseo CMОценок пока нет
- Fiebre Amarilla PDFДокумент7 страницFiebre Amarilla PDFEliseo CMОценок пока нет
- Fisiologia EndocrinaДокумент15 страницFisiologia EndocrinaEliseo CMОценок пока нет
- N0rma$ 5P5 PDFДокумент22 страницыN0rma$ 5P5 PDFEliseo CMОценок пока нет
- T6 C 2 OlogiaДокумент8 страницT6 C 2 OlogiaEliseo CMОценок пока нет
- Un1d4d de Cu1dad0s 1ntens1vosДокумент7 страницUn1d4d de Cu1dad0s 1ntens1vosEliseo CMОценок пока нет
- NUM15M4T1C4Документ8 страницNUM15M4T1C4Eliseo CMОценок пока нет
- Format Oapa General 3Документ18 страницFormat Oapa General 3Luis Enrique Persye WatzonОценок пока нет
- Fiebre Amarilla PDFДокумент7 страницFiebre Amarilla PDFEliseo CMОценок пока нет
- Sistema Respiratorio WikyДокумент10 страницSistema Respiratorio WikyJean PierreОценок пока нет
- Las Ví4s de Admini5tración de Los Med1camentosДокумент13 страницLas Ví4s de Admini5tración de Los Med1camentosEliseo CMОценок пока нет
- EpitelioДокумент7 страницEpiteliomarianaОценок пока нет
- EMBRIOLOGÍAДокумент3 страницыEMBRIOLOGÍAEliseo CMОценок пока нет
- Lectura en FríoДокумент9 страницLectura en FríoEliseo CMОценок пока нет
- Ciclo de KrebsДокумент11 страницCiclo de KrebsEliseo CMОценок пока нет
- T3cn1c4s Qu1rurg1c45Документ6 страницT3cn1c4s Qu1rurg1c45Eliseo CMОценок пока нет
- GlucólisisДокумент16 страницGlucólisisEliseo CMОценок пока нет
- D4nz45 A11t0c7on45Документ5 страницD4nz45 A11t0c7on45Eliseo CMОценок пока нет
- L11C1F3RДокумент5 страницL11C1F3REliseo CMОценок пока нет
- Pla4n3t4 m4rt3Документ4 страницыPla4n3t4 m4rt3Eliseo CMОценок пока нет
- T30r14 d3l Ju3g05Документ12 страницT30r14 d3l Ju3g05Eliseo CMОценок пока нет
- Psi 1 Qi 1 Atri 4Документ3 страницыPsi 1 Qi 1 Atri 4Eliseo CMОценок пока нет
- Técnicas Quirúrgicas EspecialesДокумент5 страницTécnicas Quirúrgicas EspecialesEliseo CMОценок пока нет
- Ob 3 Teri 1 Ci 4Документ2 страницыOb 3 Teri 1 Ci 4Eliseo CMОценок пока нет
- ANTIMICÓTICOSДокумент6 страницANTIMICÓTICOSEliseo CMОценок пока нет
- Ep 1 D 3 M 1 Ol 0 Gi 4Документ3 страницыEp 1 D 3 M 1 Ol 0 Gi 4Eliseo CMОценок пока нет
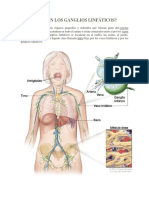
- Qué Son Los Ganglios LinfáticosДокумент10 страницQué Son Los Ganglios LinfáticosMiguel A Albites FloresОценок пока нет
- Calles de La CiudadДокумент71 страницаCalles de La CiudadEliseo CMОценок пока нет
- El Hombre Light VS El Hombre PersonalizadoДокумент5 страницEl Hombre Light VS El Hombre PersonalizadoMatias jesus Mora gonzalez100% (1)
- Trabajo Final de CriminalisticaДокумент11 страницTrabajo Final de Criminalisticacesar augusto vega caceresОценок пока нет
- Nuestra Identidad PDFДокумент146 страницNuestra Identidad PDFArnold ArnezОценок пока нет
- Hoja Resumen-Los EsponsalesДокумент2 страницыHoja Resumen-Los EsponsalesLuisaPlasenciaОценок пока нет
- Inventario de Texas RevisadoДокумент1 страницаInventario de Texas RevisadoAsistencia Tanatologica GDAОценок пока нет
- La Tradición Draconiana y El Left Hand PathДокумент5 страницLa Tradición Draconiana y El Left Hand PathDevanandaОценок пока нет
- Modelos de Correlación Entre El PDC y El CBR Inalterado PaДокумент23 страницыModelos de Correlación Entre El PDC y El CBR Inalterado PaSara Ramirez Valencia100% (2)
- Francisca Reyes Amigo Control Semana 8.Документ5 страницFrancisca Reyes Amigo Control Semana 8.Francisca Reyes AmigoОценок пока нет
- OracionesBimembresДокумент3 страницыOracionesBimembresSoledad Lopez VillavicencioОценок пока нет
- FENOMENOS DE LA MUERTE Y MEDICINA FORENSEДокумент3 страницыFENOMENOS DE LA MUERTE Y MEDICINA FORENSEJOSE FLORESОценок пока нет
- BF Skinner: Pionero del ConductismoДокумент4 страницыBF Skinner: Pionero del ConductismoCarlitos GuerraОценок пока нет
- PROYECTO Casero EstufaДокумент15 страницPROYECTO Casero EstufaGianina Luz FernandezОценок пока нет
- Contrato de servicio de verificación vehicular federalДокумент11 страницContrato de servicio de verificación vehicular federalRuben93030823Оценок пока нет
- Zapatas ProfundasДокумент8 страницZapatas Profundasmanaudi100% (1)
- Vicios de Voluntad El ErrorДокумент32 страницыVicios de Voluntad El ErrorGabo Rossel100% (1)
- Cómo Se Identifican Las Averías Por Sobretensión en TransformadoresДокумент5 страницCómo Se Identifican Las Averías Por Sobretensión en TransformadoresLedred1Оценок пока нет
- Presentacion PPT - Ruido, Iluminación y VentilaciónДокумент35 страницPresentacion PPT - Ruido, Iluminación y VentilaciónZoe MtОценок пока нет
- Cartel CientificoДокумент1 страницаCartel CientificoTéllez Nolasco Laisha JohanaОценок пока нет
- Guia Docente Latin EclesiasticoДокумент5 страницGuia Docente Latin EclesiasticoAngelis Et ArchangelisОценок пока нет
- Cómo Funciona El ProtoboardДокумент3 страницыCómo Funciona El ProtoboardJonathan CoyagoОценок пока нет
- 2 FCuantica ACTДокумент9 страниц2 FCuantica ACTrocio sanchezОценок пока нет
- Negociacion Empresarial U2 Fases Toma Decisiones PDFДокумент29 страницNegociacion Empresarial U2 Fases Toma Decisiones PDFMirgen MirandaОценок пока нет
- OS136Документ2 страницыOS136komodiemoОценок пока нет
- Cartel DHДокумент1 страницаCartel DHCarlos HerreraОценок пока нет
- Manual de Catalogación, Clasificacion - NazdiaДокумент66 страницManual de Catalogación, Clasificacion - NazdiaNazdia Yadzia Herrera MendezОценок пока нет
- Informe Final Snap Argpaz Uis 111Документ25 страницInforme Final Snap Argpaz Uis 111Johan Steven Pinto AcostaОценок пока нет
- Administracion de Datos de PozosДокумент4 страницыAdministracion de Datos de PozosUzziel De jesus OsorioОценок пока нет
- Tipos de BielaДокумент33 страницыTipos de BielaXayitho Arcos100% (7)
- Patologia GeneralДокумент24 страницыPatologia GeneralAlexLanda0% (1)
- Charlas de 5 MinutosДокумент12 страницCharlas de 5 MinutosALADINHEОценок пока нет